

{kind=link}
{kind=link}
Drought Stress Impairs Grain Yield and Quality of Rice Genotypes by Impaired Photosynthetic Attributes and K Nutrition
Cite this Article
Muhammad Zahid Mumtaz, Muhammad Saqib, Ghulam Abbas, Javaid Akhtar, Zia Ul-Qamar. 2020, Drought Stress Impairs Grain Yield and Quality of Rice Genotypes by Impaired Photosynthetic Attributes and K Nutrition. Rice Science 水稻科学(英文版), 27(1): 5-9. 
Doi:10.1016/j.rsci.2019.12.001
Permissions
Doi:10.1016/j.rsci.2019.12.001
Copyright©2020, Editorial Office of Rice Science
China National Rice Research Institute
Drought Stress Impairs Grain Yield and Quality of Rice Genotypes by Impaired Photosynthetic Attributes and K Nutrition
Drought is one of the most prevalent abiotic stresses that adversely affect rice productivity (Petrozza et al, 2014). Rice is very sensitive to drought stress and drought can cause 50% reduction in rice production globally (Yang et al, 2008). To meet the food needs for global population, 63% more agricultural production will be required by the year 2050 than the current production (Alexandratos and Bruinsma, 2012; FAO, 2017). Drought stress delays the time of flowering, leading to a reduced number of panicles, number of kernels and ultimately grain yield (Pantuwan et al, 2002; Iseki et al, 2014). The reproductive stage of rice is very sensitive to drought stress, and water limitation at this stage causes a serious reduction in rice yield (Aydinsakir et al, 2013) and biomass (Iseki et al, 2014). It is a major challenge to increase rice production under increasing drought due to changing climate, however, it may be achieved through use of drought-tolerant rice varieties having the ability to produce high yield under drought stress conditions (Luo, 2010).
This study was conducted under a split plot design with three replications at the Research Farm of the Institute of Soil and Environmental Sciences, University of Agriculture Faisalabad, Pakistan. Nursery of 11 rice genotypes (99404, 99417, Super Basmati, KS-282, KSK-434, KSK-133, Basmati-2000, KS-432, Basmati-515, Basmati-385 and Shaheen Basmati) was grown under well-irrigated conditions. Data of rainfall, sunshine, and average temperatures are given in Table 1. A well-irrigated control (non-stressed) was maintained by continuous recommended irrigations from transplanting to a week before maturity. The drought stress treatment was started at 60 d after transplantation and thereafter this field was not irrigated. There was 41.17% less application of water in case of drought treatment as compared to the well-irrigated treatment.
| Table 1 Rainfall, solar radiation and average minimum and maximum temperatures for the crop season at the experimental site. |
Drought stress affects gas exchange attributes like photosynthetic rate, stomatal conductance and transpiration rate (Serraj et al, 2011). After four weeks of drought stress, the photosynthetic rate, transpiration rate and stomatal conductance of expanded second top leaves were measured according to Mumtaz et al (2018). Drought stress caused a significant reduction in different gas exchange attributes including photosynthetic rate, transpiration rate and stomatal conductance, and significant genotypic variations were observed among different rice genotypes regarding these parameters (Table 2). KS-282 showed significantly higher photosynthetic rate, transpiration rate and stomatal conductance under drought stress as compared to the other genotypes. Drought stress caused 19%, 48%, and 47% decreases in photosynthetic rate, transpiration rate and stomatal conductance of KS-282 over the control, respectively. Meanwhile, 99404 followed by 99417 showed lower photosynthetic rate, transpiration rate and stomatal conductance with 30%, 67% and 68% decreases, respectively, as compared to control. Closure of stomata under drought stress to conserve moisture content may cause a decrease in stomatal conductance (Iseki et al, 2014). Stomatal conductance is associated with turgidity of leaves. Under drought stress, reduction in stomatal conductance also causes decreases in photosynthetic rate and transpiration rate (Rauf et al, 2015). It has been reported that stomatal conductance plays an important role in regulating the water balance of plants (Sinclair et al, 2010). If stomata are closed, the expansion of cell is reduced which leads to limited growth rate, biomass and yield production. Under severe drought conditions, CO2 fixation is reduced due to the inhibition of the activities of some key enzymes such as rubisco activase and ribulose-1, 5-bisphosphate carboxylase/oxygenase. Drought induced inhibition of enzyme activities results in the generation of reactive oxygen species, which causes photo-oxidation and harm to the photosynthetic membrane proteins, pigments and lipids, and ultimately affects grain quality (Rauf et al, 2015). Drought induced photosynthetic limitations during reproduction phase causes reduction in carbon flux to reproductive organs, triggers ovary abortion, increases pollen sterility, which leads to decreased grain yield and quality (Boyer and Westgate, 2004; Centritto et al, 2009). The results of present work suggested that drought tolerant genotypes have a better ability to maintain their physiological functions under drought stress.
| Table 2 Effects of drought stress on photosynthetic rate, transpirational rate and stomatal conductance of different rice genotypes. |
Number of panicles per plant was recorded manually before harvesting. Paddy yield was recorded after air drying the rice grains. One week sun-dried rice straw was weighed and straw yield was noted. Drought stress significantly reduced grain and straw yields as well as number of panicles per plant (Table 3). Under drought stress, KS-282 showed the highest number of panicles per plant with 7% decrease as compared to the control. The lowest number of panicles per plant was shown by 99404 with 29% decrease over the control. KS-282 also differed significantly regarding number of panicles per plant from all the other genotypes whereas 99404 was statistical at par with 99417. Genotypic variations were also observed with regard to straw and grain yields. KS-282 showed the maximum grain yield (38% decrease over the control) but was statistical at par with KS-432, Basmati-385 and Shaheen Basmati under drought stress, whereas 99404 showed the minimum grain yield with 58% decrease over the control. Similarly, KS-282 also produced the highest straw yield (21% decrease over the control) with a significant difference from all the other genotypes under drought conditions. However, the lowest straw yield (33% decrease over the control) under drought conditions was observed in 99404. These findings are supported by Pantuwan et al (2002), Yue et al (2006), Kumar et al (2009), Luo (2010) and Iseki et al (2014). Pantuwan et al (2002) tested 128 genotypes under mild to prolonged severe drought stresses at the grain filling stage and reported flowering time as an important cause of grain yield loss. Reduction in grain yield under drought stress could be due to increased spikelet sterility that reduces fertile panicles and grain weight (Yue et al, 2006). Straw yield is closely related to photosynthesis which is severely reduced under drought stress (Iseki et al, 2014).
| Table 3 Effects of drought stress on number of panicles per plant, grain yield and straw yield of different rice genotypes. |
Plants can maintain the uptake of cations to cope with drought stress (Cakmak, 2005). K+ is an important cation in plants, which plays a role in protein synthesis, membrane permeability, cell expansion, enzyme activity and stomatal opening and closing (Hopkins and Huner, 2004). K+ content in plants is positively related to transpiration rate. Drought stress accumulation with K+ deficiency causes an increase in reactive oxygen species (ROS) that induced disturbances in stomatal opening and photosynthesis (Mengel and Kirkby, 2001). Severe drought stress increases the demand for K+ to avoid oxidative damage and to protect chloroplasts. Plants also need K+ under drought stress to maintain photosynthetic CO2 fixation, which is reduced as a result of stomatal closure (Egilla et al, 2005).
Determination of K+ concentration in straw and grains were conducted by digesting the samples as described by Wolf (1982). There was a significant effect of drought stress on K+ concentration in grains and straw with a significant variation among different rice genotypes (Table 4). Significantly higher K+ concentrations in straw and grains were observed in KS-282 (20% and 36% decreases, respectively) followed by KS-432 (12% and 25% decreases, respectively) as compared to the control. The lower K+ concentrations of straw and grains were shown by 99404 as compared to the rest of the genotypes. This genotype showed 34% and 17% reductions in K+ concentrations of straw and grains, respectively, over the control. K+ content in drought tolerant genotypes is positively related to transpiration rate, however, K+ uptake may vary due to its availability in the soil. Wang et al (2013) concluded that increased K+ availability under drought stress improves enzyme activity, cell expansion, stomatal conductance, leaf area index, water use efficiency and nutrient uptake, which may promote dry matter and grain yield production. Wang et al (2004) explored the effects of K application on plant K+ uptake, grain yield and quality of japonica rice, and found that with increasing K application, plant K+ uptake is also increased, which considerably increases the number of ear bearing tillers and seed-setting rate. Moreover, high K+ content in plants is correlated with a corresponding decrease in chalkiness and amylose content and high grain yield and quality.
| Table 4 Effects of drought stress on K+ concentrations in straw and grains of different rice genotypes. mmol/g |
Grain quality is important for the acceptance and adaptation of any new cultivars by farmers and consumers (Cooper et al, 2008). Generally, in the Indian subcontinent, people prefer rice grain having medium to long grains. Market price can be estimated by grain size and shape, proportion of chalky and broken rice grain (Cooper et al, 2008). Previous studies regarding drought stress have been restricted to physiological and yield attributes but their effects on grain quality are not well-known. Three genotypes (Super Basmati, KS-282 and 99404) were selected for the determination of grain quality as affected by drought stress. Drought stress caused significant reduction in grain length and width of all the selected rice genotypes (Fig. 1). Under drought conditions, the maximum grain length was showed in Super Basmati (7% reduction) and the minimum grain length (2% reduction) was in KS-282 over control. On the contrast, KS-282 produced the maximum grain width with 6% reduction under drought stress as compared to the control. It was revealed that drought stress significantly increased the broken fraction and reduced the total milling recovery (TMR) of all the three rice genotypes (Fig. 2). The maximum broken fraction was observed in Super Basmati, and KS-282 showed the minimum broken fraction with 25% and 54% increases, respectively, under drought stress as compared to the control. Super Basmati showed the highest TMR with 4% decrease under drought stress whereas the minimum TMR was observed in 99404 with 4% reduction over the control. There were no chalkiness spots observed in any genotype under normal conditions whereas under drought stress, 9, 5 and 5 chalkiness spots were observed on grains of 99404, Super Basmati and KS-282, respectively (Table 5). We measured a negative relation between the grain length and width and these results are in accordance with the findings of Koutroubas et al (2004). Reduction in grain width could be linked with a decrease in average endosperm cell area or with abnormal amyloplast packaging that results in white chalky areas (Ishimaru et al, 2009). Fabre et al (2005) reported that grain dimensions are reduced under stress conditions. These results are also similar to the outcomes of Rao et al (2013). The results of this study also revealed a reduction in the total milling recovery along with an increase in broken fraction of rice grains due to drought stress. Super Basmati showed the minimum total milling recovery with the highest broken fraction and longer grain length. During the milling process, breakdown of longer grains was more serious than that of the shorter to medium grains. There was an inverse relationship between grain length and total milling recovery in this study. Sharifi et al (2009) also reported an interaction between rice genotype and environment with respect to grain length and shape. Similar findings have also been discussed by Adu- Kwarteng et al (2003) and Rao et al (2013).
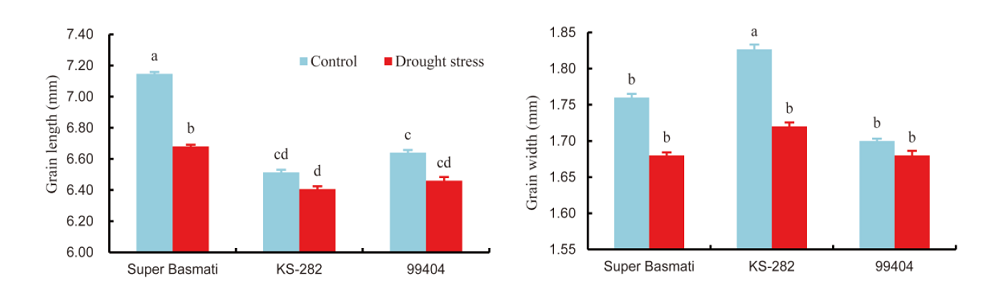 | Fig. 1. Effects of drought stress on grain length and grain width of rice genotypes. Data are Mean ± SE (n = 3). Different letters above the bar indicate significant difference according to the least significant difference test at the 0.05 level. |
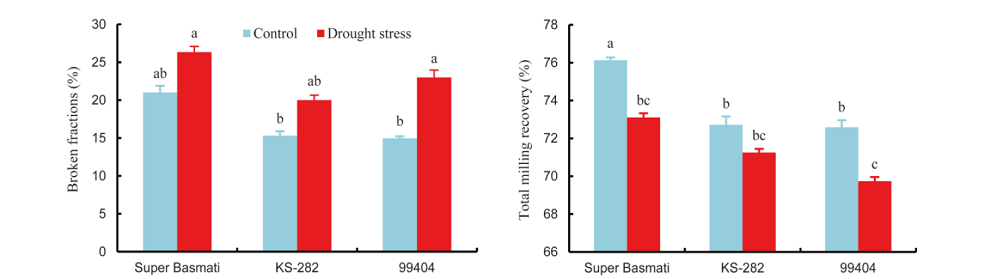 | Fig. 2. Effects of drought stress on broken fraction and total milling recovery of rice genotypes. Data are Mean ± SE (n = 3). Different letters above the bar indicate significant difference according to the least significant difference test at the 0.05 level. |
| Table 5 Effects of drought stress on chalkiness score on grains of different rice genotypes. |
Chalkiness is an important quality characteristic in the rice grain occurs commonly with the development of numerous air spaces between loosely packed starch granules and environmental stress during grain development (Tashiro and Wardlaw, 1991). In the present study, 99404 showed the greater chalkiness score under drought stress as compared to Super Basmati and KS-282. It could be due to environmental stress (drought in this case) that hindered the normal grain filling (Adu-Kwarteng et al, 2003). Under stress conditions, chalkiness in the grains causes the breakage of grains that fetch a lower price in the market. Chalky spot on grains appears as results of reduced water supply under drought stress. It is particularly evident for 99404 which is a drought-sensitive genotype with greater chalkiness score. Chalkiness degrades the rice appearance, transparency, head rice recovery and consumer acceptability (Graham, 2002; Yoshioka et al, 2007). It is negatively correlated with milling quality as increase in chalkiness caused decrease in head rice recovery (Wassmann et al, 2009; Zhao and Fitzgerald, 2013).
SUPPLEMENTAL DATA
The following material is available in the online version of this article at http://www.sciencedirect.com/science/journal/ 16726308; http://www.ricescience.org.
Supplemental File 1. Materials and methods used in this study.
Reference
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|