{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
MULTI-FLORET SPIKELET 4( MFS4) Regulates Spikelet Development and Grain Size in Rice
[Wang Yan# , Zeng Xiaoqin# , Lu Lu, Cheng Qinglan, Yang Fayu, Huang Mingjiang, Xiong Mao, Li Yunfeng ]
]
]
|
|


#These authors contributed equally to this work
In rice, the spikelet is the basic unit of inflorescence, and its development is important for determining the grain yield and quality. We reported a rice spikelet mutant multi-floret spikelet 4 ( mfs4) which resulted in the production of extra floral organs or a whole extra floret, and elongated sterile lemmas. The results suggested that the mutation of the MFS4 gene interfered with spikelet meristem determinacy and floral organ identity. In addition, the plant height and the grain length and width in the mfs4 mutant were all less than those in the wild type. Using the bulked segregant analysis method, the MFS4 gene was localized in a 557-kb region on the long arm of chromosome 1. Sequence analysis showed that there was a C-base deletion at the open reading frame of LOC_Os01g67430. Further tests indicated that a wild type copy of LOC_Os01g67430 was able to reverse the mfs4 defects, which indicated that LOC_Os01g67430 was the MFS4 gene. The MFS4gene encodes a lipase located in the mitochondria and is expressed strongly in the young inflorescence. qRT-PCR results showed that the expression of some genes that were known to regulate spikelet meristem determinacy and grain size were decreased in the mfs4 mutant, which indicated that the MFS4 gene regulates spikelet meristem determinacy and grain size by modulating the expression of these genes.
Rice (Oryza sativaL.) is one of the world’ s most important food crops. Its yield is determined by three key factors: grain number per panicle, seed-setting rate and 1000-grain weight. To optimize these parameters, it is crucial to understand the genetic and molecular mechanisms of the development of the panicle, spikelet and grain. It is well known that the number and length of the branches determine the grain number per panicle to a large degree. Many genes, such as IDEAL PLANT ARCHITECTURE(IPA1), CYTOKININ OXIDASE/DEHYDROGENASE(OsCKX2), DENSE AND ERECT PANICLE 1 (DEP1) and GRAINS HEIGHT DATE-7(Ghd7), are involved in the regulation of initiation and growth of the branch meristem and affect the number of spikelets per grain, and have been identified as yield-related factors (Ashikari et al, 2005; Xue et al, 2008; Huang et al, 2009; Zhang et al, 2017). The spikelet is a basic and unique inflorescence structure in grass, which consists of a pair of bracts and 1‒40 florets per grain (Malcomber et al, 2006). However, few researchers have considered if and how the number of florets per grain within the spikelet affects the grain number per panicle or the yield.
As with the inflorescence, the spikelet can be classified as determinate or indeterminate according to whether the apical meristem transforms into a terminal flower meristem (FM). In determinate spikelet species such as rice and maize, the spikelet meristem (SM) is terminated by the formation of the terminal FM after developing a fixed number of lateral FMs. In contrast, in indeterminate spikelet species such as wheat, there are no terminal FMs, thus the SM cannot be terminated and always maintains the ability to form an indeterminate number of lateral FMs (Malcomber et al, 2006). Several genes that encode the AP2/ERF domain have been reported to regulate SM fate determinacy, for example, INDETERMINATE SPIKELET 1 (IDS1) in maize (Chuck et al, 2008), and SUPERNUMERARY BRACT (SNB) and OsIDS1 in rice (Lee et al, 2007; Lee and An, 2012; Ren et al, 2013). The Ids1 mutant of maize produces an extra floret within the spikelet, which suggests that the IDS1 gene is involved in maintaining SM determinacy (Chuck et al, 2008). Both SNB and OsIDS1 are orthologs of the IDS1 gene in rice. A mutation of SNB results in the production of extra rudimentary glumes or florets (Lee et al, 2007). Although there are no obvious defects in osids1 single mutants, and minimal defects in snb single mutants, the snb+osids1 double mutant shows a more severe phenotype than the single mutants (Lee and An, 2012). In addition, the MULTI-FLORET SPIKELET 1 (MFS1) gene, another AP2/ERF transcription factor, can also promote SM-to-FM transformation by activating the SNB and OsIDS1 genes in rice (Ren et al, 2013). These findings suggest that these AP2-like AP2/ERF factors may initiate SM-to-FM transformation before terminating the SM. These AP2-like genes are targets of microRNA172s (miR172s). The suppression of their expression by miR172s is necessary for SM determinacy (Zhu et al, 2009; Lee and An, 2012). In addition, EXTRA GLUME 1 (EG1)/DOUBLE-FLORET 1 (DF1), DEPRESSED PALEA 1 (DP1)/MULTI-FLORET SPIKELET 3(MFS3) and MULTI-FLORET SPIKELET 2 (MFS2)/MORE FLORET 1 (MOF1) encode lipase, AT-hook protein and MYB protein, respectively, as well as regulating spikelet determinacy in rice. Loss of function of these genes results in the formation of two florets or extra glumes in spikelets, which indicates that the mutation of these genes may enable the transformation of the meristems from determinate to indeterminate (Li et al, 2009; Jin et al, 2011; Ren et al, 2018b, 2020; Zheng et al, 2019; Li et al, 2020).
In the present study, we identified a rice MULTI- FLORET SPIKELET 4 (MFS4) gene, which encodes a lipase and is located in the mitochondria. The mfs4 mutant generated extra floral organs or a whole extra floret, which suggested delayed SM-to-FM transformation. In addition, decreased grain length and width in the mfs4 mutant indicated that MFS4 also regulated the development of the grain size. Our results showed that the MFS4 gene regulated the determinacy of SM by affecting the expression of AP2-like genes, and regulated grain size by modulating the expression of related genes.
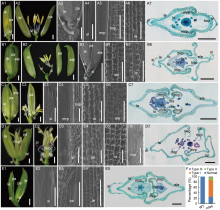
The wild type (WT) spikelet was composed of a pair of rudimentary glumes, a pair of sterile lemmas and a fertile terminal floret. The WT floret was composed of four whorls of floral organs from the outside to the inside. The lemma and the palea (whorl 1) were closely coupled, two lodicules (whorl 2) were located on the lemma side, and six stamens (whorl 3) surrounded a pistil (whorl 4) with two stigmas (Fig. 1-A1 and -A2). The lemma and the palea, collectively termed the hulls, hooked together, and enclosed the inner floral organs. The sterile lemma and rudimentary glumes were formed in a 1/2 alternate arrangement with the hulls (Fig. 1-A3). The sterile lemma had a smooth abaxial surface with a few trichomes (Fig. 1-A4). The palea can be considered a combinational organ consisting of two parts: the body of the palea (bop) and the marginal region of palea (mrp). The bop was anatomically similar to the lemma, and the four cell layers (silicifi ed abaxial epidermis, fi brous sclerenchyma, spongy parenchymatous cells and non- silicifi ed adaxial epidermis) were discernible. The mrp consisted of a smooth non-silicifi ed epidermis, a large amount of spongy parenchymatous tissues and a small number of sclerenchyma cells (Fig. 1-A5 to -A7; Fig. 2-A and -B). The lemma consisted of a silicified abaxial epidermis and bore trichomes and protrusions (Fig. 1-A6). The lemma contained five vascular bundles, the palea contained three vascular bundles and the sterile lemma contained one vascular bundle (Fig. 1-A7 and Fig. 2-C).
 | Fig. 1. Phenotypes of spikelet in wild type (WT) and mfs4 mutants. A, Spikelet of the WT. A1, WT spikelet; A2, The lemma and palea were removed in A1; A3 to A6, The WT spikelet surface characters of sl, pa and le; A7, Transverse section of WT spikelet. B‒E, Spikelets of the mfs4 mutants. B1 and B2, The Type I mfs4mutant spikelet; B3‒B5, The Type Imfs4 mutant spikelet surface characters of spikelet, le and ele; B6, Transverse section of the Type Imfs4 mutant. C1 and C2, The Type IImfs4mutant spikelet; C2, The lemma and extra lemma were removed; C3‒C6, The Type IImfs4 mutant spikelet surface characters of sl, pa, le and ele; C7, Transverse section of the Type IImfs4 mutant. D1 and D2, The Type IImfs4 mutant spikelet; D2, The lemma, palea and elongate sterile lemma were removed; D3‒D6, The Type IImfs4 mutant spikelet surface characters of sl, pa, le and mrp; D7, Transverse section of the Type IImfs4 mutant. E1, The Type III mfs4 mutant spikelet; E2‒E4, The Type IIImfs4 mutant spikelet surface characters of sl, pa and le; E5, Transverse section of the Type IIImfs4 mutant. F, Percentage of mutant organs in mfs4 spikelets. |
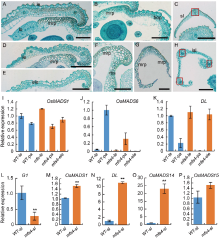
 | Fig. 2. Histological and qRT-PCR analysis of lateral organs in spikelets of wild type (WT) and mfs4 mutants. A‒C, Transverse sections of lateral organs in the WT spikelet. A and B, The lemma and palea of WT. C, The sterile lemma of WT. Red box indicates the vascular bundle. Scale bars are 500 μ m. D‒H, Transverse sections of lateral organs in the mfs4 mutant spikelets. D‒G, The lemma, palea and extra lemma of the mfs4 mutant spikelets. H, The elongate sterile lemma of the mfs4 mutant spikelets. Red boxes indicate the vascular bundles. Scale bars are 500 μ m. I‒P, qRT-PCR analysis of OsMADS1, OsMADS6, DL, G1, OsMADS14 and OsMADS15expression. Actin was used as a control. RNA was isolated from the flower organs of the WT and mfs4mutant spikelets. Data are Mean ± SE (n = 3). * , P ≤ 0.05 and * * , P ≤ 0.01 by the Student’ s t-test. bop, Body of palea; ele, Extra lemma; le, Lemma; lo, Lodicule; lsl, Lemma- like sterile lemma; mrp, Marginal region of palea; Pa, Palea; bs, Residual bop-like structure; sl, Sterile lemma. bop, Body of palea; ele, Extra lemma; le, Lemma; lo, Lodicule; mrp, Marginal region of palea; lsl, Lemma-like sterile lemma; ov, Ovule; pa, Palea; pi, Pistil; rbs, Residual bop-like structure; rg, Rudimentary glume; sl, Sterile lemma; st, Stamen. Scale bars are 1 000 μ m in A1, A2, B1, B2, C1, C2, D1, D2 and E1; Scale bars are 500 μ m in A3‒A6, B3‒B5, C3‒C6, D3‒D6 and E2‒E4; Scale bars are 100 μ m in A7, B6, C7, D7 and E5. |
Most significantly, the mfs4 mutant spikelets produced extra floral organs or occasionally a whole additional floret. Three types of phenotypes with indeterminate spikelets were found in the mfs4 mutant from the statistical analysis. In the Type I spikelets (62% of mfs4 spikelets), an additional lemma-like organ and a whole fertile terminal floret were formed on the spikelet rachilla (Fig. 1-B1 to -B6, -F and Fig. 2-E). In the Type II mutant spikelets (11% of mfs4 spikelets), instead of the terminal floret, two incomplete florets were formed. In one Type II mutant spikelet, in addition to rudimentary glumes and sterile lemmas, the spikelet consisted of two lemma-like organs, two degraded paleas (each one consisted of one residual bop-like structure and two normal mrps, or two independent mrp-like structures without a bop), two pairs of lodicules, 9‒12 stamens and 1‒2 pistils (Fig. 1-C1 to -D7, -F and Fig. 2-D and -G). In the Type III mutant spikelets (2% of mfs4 spikelets), two almost complete and independent florets developed, although the identity and the number of stamens in some lower florets showed differences from the WT (Fig. 1-E1 to -E5 and -F).
To further clarify the identities of the extra floral organs in the mfs4 mutant, the expression of several genes for the lemma and/or the palea identity (OsMADS1 for the lemma and the palea, OsMADS6 for the palea and DL for the lemma; Jeon et al, 2000; Nagasawa et al, 2003; Li et al, 2010) was investigated. In WT, while OsMADS1 was expressed in both the lemmas and the paleas, OsMADS6 was highly expressed in the paleas and lowly in the lemmas. Conversely, DL was expressed highly in the lemmas and weakly in the paleas. In the extra lemma-like organs of the mfs4 spikelets, the expression levels of these genes were similar to those in the WT lemmas and the upper floret-lemmas of the mfs4 spikelets (Fig. 2-I to -K). This indicated that these additional lemma-like organs were just lemmas. In the degraded paleas of the mfs4 mutant spikelets, the transcription levels of DL and OsMADS1 were similar to those in the WT paleas, whereas the transcription level of OsMADS6 was down- regulated significantly in the mfs4 mutants (Fig. 2-I to -K). This result suggested that these palea-like organs were indeed paleas and consistent with the phenotype of the mfs4 mutants.
The sterile lemma was elongated or transformed into a lemma-like organ in some mfs4mutant spikelets. The WT sterile lemma had a relatively smooth upper epidermis bearing a few trichomes (Fig. 1-A4). However, the lemma-like organ was similar to the sterile lemma in size and shape. Scanning electron microscope (SEM) images showed that the upper epidermis of the elongated mfs4 mutant sterile lemma developed many trichomes and silicifi ed cells similar to those in the upper epidermis of the WT lemma (Fig. 1-D3). Histological analysis also showed that the WT sterile lemma only contained one vascular bundle (Fig. 2-C), but the mfs4 mutant sterile lemma contained three vascular bundles (Fig. 2-H). We then detected the expression levels of G1(a sterile lemma identity gene), and OsMADS1, OsMADS14, OsMADS15 andDL(four lemma/palea identity genes) in the mfs4 mutant elongated sterile lemmas (Nagasawa et al, 2003; Arora et al, 2007; Yoshida et al, 2009; Wang et al, 2010). The qRT-PCR results indicated that three lemma/palea identity genes (OsMADS1, OsMADS14 and DL) were up-regulated significantly in the elongated sterile lemmas of the mfs4 mutants, while the sterile lemma identity gene G1 was down- regulated sharply (Fig. 2-L to -O). These results indicated that the elongated sterile lemmas in the mfs4 mutant spikelets had gained lemma-like identities to a large extent.
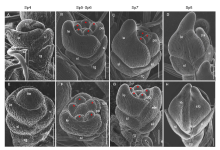
The early development of rice spikelets can be divided into stages Sp1‒Sp8 (Ikeda et al, 2004). SEM was used to observe the difference between the WT and mfs4mutant spikelets at the early stage. In the WT spikelets, the palea primordia were initiated at Sp4 (Fig. 3-A). Then, two pairs of lodicule primordia formed at Sp5 were covered by the lemma and palea. At Sp6, six stamen primordia formed, and a carpel primordium formed at Sp6‒Sp7. During the Sp8 stage, the primordia of lemma palea were hooked together and enclosed the inner floral organs (Fig. 3-B to -D). In the mfs4 mutant, the development of floral organ primordia differed from the WT. From the Sp5 stage, extra lemma primordia between the lemma and the sterile lemma primordia were observed and the extra lemma primordia progressed in a manner/sharp size similar to that of the lemma primordia (Fig. 3-E to -H). These results showed that the defects in the mfs4mutant spikelets were occurred during the early developmental stages.
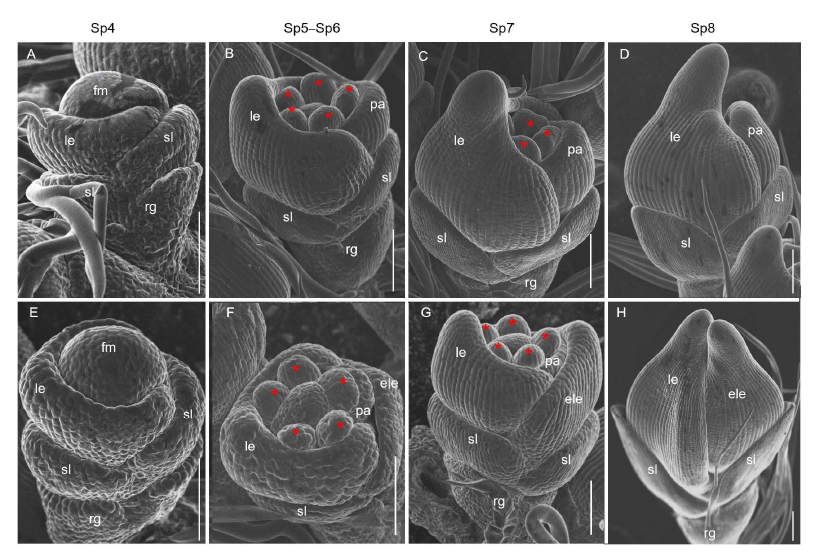 | Fig. 3. Scanning electron micrographs of spikelets at early developmental stages in wild type (WT) and mfs4 mutant. A-D, WT. E-H, mfs4 with ele. Asterisks indicate the stamens. Scale bars are 100 μ m. ele, Extra lemma; fm, Flower meristem; le, Lemma; pa, Palea; rg, Rudimentary glume; sl, Sterile lemma. |
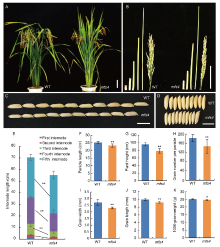
In addition to the spikelet defects, we noted that the mfs4 mutants showed some changes in the plant and grain. Firstly, the plant height of the mfs4 mutant was significantly shorter than that of the WT (Fig. 4-A and -G). In the WT plants, five nodes were elongated, while only four elongated nodes were observed in the mfs4 plants (Fig. 4-B). Furthermore, the first three internodes and the panicle length of the mfs4 mutant were significantly shorter than those of the WT (Fig. 4-E and -F), which indicated that MFS4 affected stem development in rice. Secondly, the grains of the mfs4 mutants were significantly shorter and narrower than those of the WT, which resulted in a decrease in 1000-grain weight (Fig. 4-I to -K). Finally, there was significantly fewer grain number per panicle in the mfs4 mutants than that in the WT (Fig. 4-H). These results showed that MFS4 affected the number and size of the grains, and hence the yield in rice.
 | Fig. 4. Comparison of morphological characters between wild type (WT) and mfs4 mutant. A, Plant morphology of WT and mfs4 mutant at the heading stage. B, Internodes and panicles of WT and mfs4 mutant. C and D, Comparisons of grain length (C) and width (D) in the WT and mfs4 mutant. E‒K, Comparisons of internode length (E), panicle length (F), plant height (G), grain number per panicle (H), grain width (I), grain length (J) and 1000-grain weight (K) between the WT and mfs4 mutant. Scale bars are 10 cm in A and B, and 1 cm in C and D. * , P ≤ 0.05 and * * , P ≤ 0.01 by the Student’ s t-test. |
To isolate the MFS4 gene, the mfs4 mutant was crossed with a sterile line 56S (indica). All F1 plants manifested the WT phenotype. F2 population resulted in 1 080 plants in total, among them, 831 plants were normal and 249 plants manifested the mfs4 mutant phenotype. The segregation ratio of WT to mutant plants was conformed to 3:1 (χ 2 = 3.11 < χ 20.05 = 3.84), which indicated that the multi-floret spikelet trait is controlled by a single recessive gene.
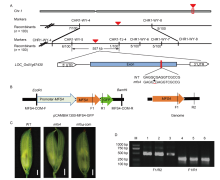
Recessive individuals in the F2 population were used as a mapping population to localize the MFS4 gene. Within the F2 segregating population, 15 WT plants and 15 mutant plants were selected randomly to construct WT and mutant DNA pools, respectively. Linkage analysis showed that the MFS4gene was linked to the polymorphic SSR markers CHR1-WY-4, CHR1-TJ-4, CHR1-WY-5, CHR1-WY-6, CHR1-WY-7 and CHR1-WY-8 on chromosome 1. We used these markers to survey 100 mutant individuals, and the MFS4 gene was further localized in the 557 kb region between CHR1-WY-5 and CHR1-TJ-4 on the long arm of chromosome 1. Through genome re-sequencing and general PCR sequence analysis, we found a C-base deletion in the open reading frame of LOC_ Os01g67430 at the 1102th base. This deletion caused a frame shift and the premature termination of translation (Fig. 5-A). Next, we constructed a complementary vector, which contained 2000-bp upstream sequence and the full genomic sequence of LOC_Os01g67430, and then introduced the recombinant plasmid into the mfs4mutant. In total, 15 positive transgenic lines were obtained. In eight of these, themfs4 mutated traits were rescued by designed MFS4 endogenous and exogenous primers via PCR verification (Fig. 5-B to -D). The results verified that LOC_Os01g67430 is the MFS4gene.
 | Fig. 5. Fine mapping and sequencing analysis of MFS4 gene. A, Gene mapping of MFS4 on rice chromosome 1. The red triangles indicate the target gene. The red line in the exon indicates the mutational site of candidate gene. WT, Wild type. B, Schematic structure of the complementary vector pCAMBIA1300-MFS4-GFP and MFS4 genome. MFS4-COM-F/MFS4-COM-R are complementary vector primers, F1/R2 are primers for amplifying endogenousMFS4, and F1/R1 are primers for amplifying exogenousMFS4-GFP. C, Defect in mfs4 flowers were completely rescued by introduction of pCAMBIA1300-MFS4. mfs4-com, mfs4complementary flower. Scale bars are 1 mm. D, Identification of transgenic plants. M, DNA marker; Lanes 1‒3, Detection of positive transgenic plants by F1/R2 primers; Lanes 4‒6, Detection of positive transgenic plants by F1/R1 primers. |
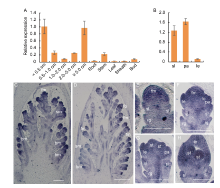
To further understand the functions of the MFS4gene, we examined its spatial expression pattern by qRT- PCR. In the vegetative tissues, MFS4 had an obvious expression in the stem, while its expression levels were significantly lower in other vegetative tissues (root, leaf, sheath and bud) (Fig. 6-A). In the reproductive tissues, MFS4 was expressed highly in young panicles and decreased gradually in inflorescences with different lengths. At the reproductive growth stage, MFS4 was expressed mainly in the young panicle and spikelet organs, and especially highly in the sterile lemmas and paleas (Fig. 6-B). Further, we detected the expression pattern of MFS4 using in situ hybridization and found strong signals in the rice panicles (Fig. 6-C to -H). The MFS4 transcripts were firstly detected in the branch meristem, spikelet meristem and floral meristem (Fig. 6-C). At the Sp3 stage when the primordia of the rudimentary glume and the sterile lemma had been initiated, the MFS4 gene was highly expressed in these organs, but not in the spikelet meristem (Fig. 6-D and -E). After the Sp4 stage, when the primordia of the floral organs were initiated, a strong MFS4 signal was detected in the primordia of floral organs, including the lemma, palea, lodicule, stamen and floral meristem. Simultaneously, MFS4 was still expressed obviously in the rudimentary glume and sterile lemma (Fig. 6-F to -H). These results indicated that MFS4 was mainly expressed in the spikelets and florets, consistent with its role in influencing spikelet meristem development and grain size.
 | Fig. 6. Spatiotemporal expression pattern of MFS4 gene. A and B, qRT-PCR of MFS4. Actin was used as a control. RNA was isolated from young panicles < 0.5 cm, 0.5‒1.0 cm, 1.0‒2.0 cm, 2.0‒5.0 cm and ≥ 5.0 cm, as well as vegetative organ and floral organ of wild type plants. Data are Mean ± SD (n = 3). C‒H, In situhybridization expression in panicles of wild type (C and D), and spikelets at Sp2‒Sp3 (E), Sp4‒Sp5 (F), Sp5‒Sp6 (G) and Sp7‒Sp8 (H). bm, Branch meristem; fm, Floral meristem; fo, Floral organ; le, Lemma; lo, Lodicule; pa, Palea; rg, Rudimentary glume; sl, Sterile lemma; sm, Spikelet meristem; st, Stamen. Scale bars are 500 μ m in C and D, and 100 μ m in E‒H. |
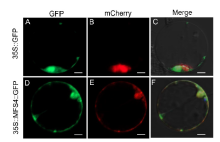
To explore the subcellular localization of the MFS4 protein, we expressed an MFS4::GFP fusion protein in rice protoplasts. We also coexpressed a mitochondrial marker MTS::Cherry fusion protein in rice protoplasts (Zhang et al, 2016). While the GFP protein alone was uniformly expressed in the cytoplasm and nucleus (Fig. 7-A to -C), it was found that the MFS4::GFP protein was co-located with the mitochondrial marker MTS::Cherry protein (Fig. 7-D to -F). These results indicated that the MFS4 protein was predominantly located in the mitochondria.
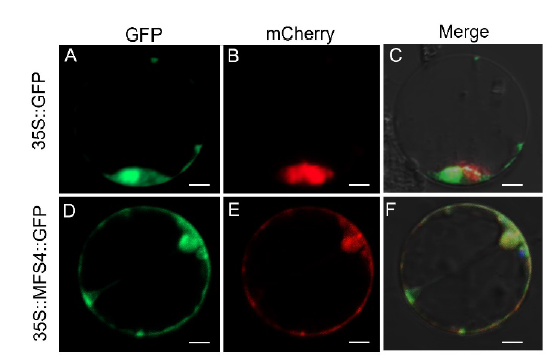 | Fig. 7. Subcellular localization of MFS4 protein. A and D, GFP illuminant. B and E, mCherry illuminant which indicates the mitochondria marker. C and F, Merge illuminant. Scale bars are 50 μ m. |
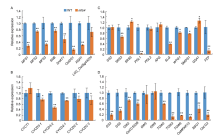
To confirm how SM development was affected in the mfs4 mutant, the expression levels of genes that regulate SM development, including LOC_Os06g43220, SNB, SHAT1, OsIDS1, RSR1, MFS1, MFS2/ MOF1 and MFS3/DP1 (Zhu et al, 2009; Huang et al, 2009; Zheng et al, 2019; Li et al, 2020; Ren et al, 2020) were detected by qRT-PCR. Compared with the WT, the expression levels of all these genes were clearly down-regulated in the mfs4 mutant (Fig. 8-A). This result indicated that MFS4 was a key upstream regulator in SM determinacy by activating AP2-like genes, MFS2/MOF1 and MFS3/DP1.
 | Fig. 8. Expression levels of related genes in wild type (WT) and mfs4 mutant. A, Expression analysis of related genes influencing spikelet meristem determination in young panicles. B, Expression analysis of related genes influencing grain size by cell cycle-related genes in panicles. C, Expression analysis of related genes influencing grain size by hull cell expansion-related genes in panicles. D, Expression analysis of related genes influencing grain size by hull cell proliferation-related genes in panicles. Actin was used as a control. RNA was isolated from panicles of WT and mfs4 mutant. At least three replicates were performed and the mean value was used. Error bars mean standard error. * , P ≤ 0.05 and * * , P ≤ 0.01 by the Student’ s t-test. |
In the mfs4 mutant panicles, around 75% of the spikelets showed defects surrounding the meristem and organ identities, but the remaining 25% showed a ‘ normal’ phenotype (Fig. 1-F). We further investigated the grain traits of these ‘ normal’ grains in the mfs4 mutants, and found that these ‘ normal’ mfs4 grains were shorter and narrower than the WT grains (Fig. 4-I and -J). To determine how MFS4 regulates grain size, we firstly investigated the expression of several genes that regulate both the cell cycle and cell expansion. Five cell cycle-related genes, CYCD3-1, CYCD3-2, CYCD2-1, CYCD2-2andCYCD2-3, showed significantly lower expression levels in the mfs4 mutant than in the WT (Fig. 8-B). The expression of CYCT1 was slightly up-regulated in the mfs4 mutant compared with WT (Fig. 8-B). We also investigated the expression of several genes that regulate grain size by hull cell proliferation and expansion. GS2, SRS3, SRS5, PGL1, PGL2, APG, SLG, WTG1, SMOS1, GSA1 and FZP influence cell expansion, and GS3, GS5, GW2, OsCCS52B, GW5, GW8, TGW2, TGW3, TGW6, OsMKKK10, BSG1, OsLG3, GSA1 and FZP are involved in regulating cell proliferation (Kitagawa et al, 2010; Heang and Sassa, 2012a; Hu et al, 2015; Ren et al, 2018a; Xiong et al, 2018; Dong et al, 2020; Guo et al, 2020). Compared with WT, the expression levels of GS3, SRS3, PGL1, GS5, GW2, OsCCS52B, SLG, WTG1, BSG1, SMOS1, FZP, TGW2, TGW3, TGW6, OsMKKK10, BSG1 and OsLG3 were reduced, while SRS5 and GSA1 were increased in the mfs4 mutant panicles. In addition, there were no obvious differences in the expression levels of GS2, APG, PGL2, GW5 and GW8 between the mfs4 and WT panicles (Fig. 8-C and -D). Together, these results supported that MFS4 affected grain size by regulating cell proliferation and expansion in the lemmas and paleas.
In this study, a spikelet-defective mfs4 mutant was identifi ed. Map-based cloning and genome DNA sequencing showed that MFS4 is a new allele of EG1/ DF1. MFS4was identified definitively as a deletion mutant allele at the EG1/DF1 locus. The genetic background of eg1-1is indica rice ZF802, in which the abnormal phenotype includes the occurrence of glume-like organs, and in some extreme cases, alteration of the number of floral organs and losses of flower determinacy (Li et al, 2009). The genetic background of eg1-2 is japonica rice ZH11, in which only the empty glumes, lodicules and stamens are affected in the abnormal phenotype (Zhang et al, 2016). The genetic background of eg1-3 is japonica rice 9522, which manifests extra glume-like structures between the sterile lemma and the lemma, together with altered floral organ numbers and identities (Cai et al, 2014). eg1-4, eg1-5andeg1-6 are derived from Nipponbare CRISPR-Cas mutations (Li et al, 2009; Zhang et al, 2016). The df1 mutant produces a ‘ two-flower’ phenotype and shows increased yield (Ren et al, 2018) b. However, the mfs4 mutant possesses the eg1/df1 allele for all phenotypes, as found in previous studies (Li et al, 2009; Cai et al, 2014; Ren et al, 2018). This suggests that the MFS4 gene might manifest different phenotypes in different genetic background materials and mutation sites. More detailed analyses of the functional alteration of the novel mutant allele mfs4 can yield new insights into the function of theEG1locus in the development of spikelet determinacy in rice and other grasses.
SM determinacy is associated with the number of lateral organs and florets produced in the spikelet. In the WT rice, because one terminal FM is formed on the top of the SM, the determinate spikelet ultimately produces one pair of rudimentary glumes, one pair of sterile lemmas and one terminal floret. Our findings revealed that the mfs4 mutant spikelets produced an extra lemma and formed two florets instead of one terminal floret. Similarly, in the mfs1, mfs2/mof1 andmfs3mutants, some spikelets produce an additional lemma-like organ or two florets (Ren et al, 2013, 2020; Zheng et al, 2019; Li et al, 2020). Moreover, in the snb mutant, some spikelets develop supernumerary rudimentary glumes, additional lemma/palea-like structures or extra florets. snb+osids1 mutants manifest a more severe phenotype than snb and osids1 single mutant (Lee et al, 2007; Lee and An, 2012). These findings suggested that MFS4, together with MFS1, MFS2, MFS3, SNB andOsIDS1, affect the SM to FM transition, and their mutations induce SM indeterminacy.
Loss of SM determinacy in these mutants tends to produce ‘ two-floret’ spikelets (Fig. 1-E) (Lee and An, 2012; Ren et al, 2013; Li et al, 2020), which suggested the possibility of a rice spikelet transforming into an indeterminate spikelet and producing two or more florets in one spikelet, similar to other indeterminate spikelet species such as wheat. If confirmed, the number of grains per panicle, and hence the rice yield, can increase significantly.
Grain size is determined by grain length, width, thickness and fullness. The development of the hulls (the lemma and the palea) is very important for grain length and width in rice and other grass crops. Many grain size regulating genes related to hull development have been identified and are widely used in rice breeding. For example, GW2 encodes a RING-type E3 ubiquitin ligase and negatively regulates cell division by anchoring its substrate to the proteasomes responsible for degradation. The loss of function of GW2 activates the division of the hull cells, thus increasing the hull width, grain width, grain weight and yield in rice (Song et al, 2007; Yan et al, 2011). GW5 regulates grain size and grain weight through a ubiquitin-proteasome pathway that is similar to GW2. The loss of function of GW5 activates the division of the hull cells, thus increasing the grain size (Wan et al, 2008). BSG1 encodes a DUF640 domain protein. In the bsg1 mutant, the number of hull cells decreases, leading to reduced grain length, grain thickness and grain weight (Li et al, 2012; Ma et al, 2013). Besides these genes, GS3 (Xia et al, 2018), GS5 (Li et al, 2011), OsCCS52B (Su’ udi et al, 2012), GW8 (Wang et al, 2012), TGW2 (Ruan et al, 2020), TGW3 (Ying et al, 2018), TGW6 (Ishimaru et al, 2013), OsMKKK10 (Ren et al, 2016), BSG1 (Li et al, 2012), OsLG3 (Xiong et al, 2018) and GSA1 (Dong et al, 2020) are also involved in regulating cell proliferation, and their mutants produce small and light grains. The grain length gene SRS3 is grouped as a member of the kinesin-13 subfamily. In the srs3 mutant, a reduction in the longitudinal cell length of the grains results in small and round seeds (Kitagawa et al, 2010; Wu et al, 2014; Deng et al, 2015). PGL1 and PGL2 are helix- loop-helix proteins that interact with APG to determine rice grain length by controlling the length of lemma/ palea cells (Heang and Sassa, 2012a, b). Genes GS2, SRS5, SLG, WTG1, SMOS1, GSA1 and FZP also influence cell expansion-regulated hull size, and their mutants display decreased hull size (Segami et al, 2012; Aya et al, 2014; Duan et al, 2016; Feng et al, 2016; Huang et al, 2017). In addition, cell cycle- related CYCD-like genes are involved in hull development by affecting cell proliferation and expansion (La et al, 2006; Ren et al, 2018a).
In the present study, we found that the grain length, grain width and 1000-grain weight of the mfs4 mutant were significantly decreased compared to the WT, and the MFS4 gene was strongly expressed in the lemma and palea. Further, in the mfs4 mutant, the expression of several genes related to hull cell proliferation and expansion (GS3, GS5, PGL1, SLG, TGW2, TGW3, TGW6, OsMKKK10, BSG1, OsLG3, CYCD3-2 andFZP) was significantly down-regulated compared to the WT. In light of all these results, we suggested that the MFS4 gene plays a crucial role in regulating the development of grain size via genes related to hull cell proliferation and expansion, and thus affecting the yield of in rice. Further understanding of the molecular mechanisms of MFS4regulating hull development could be of significance in the design of grain morphology in molecular breeding.
The mfs4 mutant was derived from the ethyl methane sulfonate (EMS) mutagenesis population of a maintainer line XD1B (indica). The mfs4 mutant was crossed with 56S (a temperature- sensitive sterile line) to generate an F1 population. In the second year, the F1population was cultivated and self-crossed to generate an F2population. All plants with the mutant phenotype in the F2population were used to locate the MFS4gene.
During the flowering stage, the spikelets of the WT and mfs4 mutants were observed and photographed using a stereoscope (SMZ1500, Nikon, Japan) and a scanning electron microscope (SU3500, Hitachi, Japan) under a -20 º C vacuum environment.
Histological analysis of the spikelets at the flowering stage was performed by paraffin section. The WT and mutant spikelets at the heading stage were immersed in FAA solution (50% anhydrous ethanol, 0.9 mol/L glacial acetic acid and 3.7% formaldehyde) and fixed at 4 º C for more than 16 h after vacuumed. Then, the spikelets were dehydrated with ethanol series and infiltrated with xylene before being embedded in paraffin (Sigma-Aldrich Inc., Shanghai, China). Next, the paraffin block that contained individual spikelets was cut into 8‒10 µ m-thick slices, which were then pasted onto microscope slides (RM2245, Leica, Hamburg, Germany). These slices were dyed sequentially with 1% safranin (Amresco Inc., Framingham, MA, USA) and 1% Fast Green (Amresco Inc., Framingham, MA, USA), then dehydrated through an ethanol series, infiltrated with xylene, and finally mounted beneath a coverslip. Light microscopy was performed using a microscope (E600, Nikon, Japan).
The mutant individuals in the F2 population were used as the mapping population to locate the target gene by the bulked segregant analysis (BSA) method. The DNA of the parents, F2 population, WT and mutant gene pools were extracted from fresh leaves following the cetyl trimethyl ammonium bromide (CTAB) method. SSR markers distributed evenly on the 12 rice chromosomes were used for gene mapping. All the primers were synthesized by the Tsingke Company (Beijing, China). The total volume of the PCR amplifications was 15 µ L, which comprised 1.5 µ L of 10× PCR buffer, 1 µ L of 50 ng/μ L DNA, 0.75 µ L of 2.5 mmol/L dNTPs, 9.5 µ L of ddH2O, 1 µ L of 10 mmol/L forward and reverse primers and 0.25 µ L of 5 U/µ L rTaq DNA polymerase. Amplification was done under the following conditions: 5 min at 94 º C for DNA strand separation, followed by 35 cycles of denaturing at 94 º C for 30 s, annealing at 56 º C for 30 s, extension at 72 º C for 30 s, and finally extension at 72 º C for 7 min. The amplified products were observed and analyzed after separated by electrophoresis on 10% polyacrylamide gels and the silver staining. The relative distances between the mfs4 locus and the linkage SSR or the insertion/deletion (InDel) markers were marked by the number of recombinants. The primers are shown in Table S1.
| Table S1. Sequences of primers for gene mapping. |
RNAs from different tissue samples of the WT and mutant were extracted using Eastep Super Total RNA Extraction Kit (Promega, China). The first-strand cDNA was synthesized from 1 µ g total RNA with oligo (dT)18 primers in a 20 µ L reaction volume using the PrimeScript® RT Reagent Kit with gDNA Eraser (TaKaRa, Dalian, China). The qRT-PCR analysis was performed with three replicates using SYBR Premix Ex Taq II Kit (Takara, Dalian, China) with an ABI 7500 Sequence Detection System (Applied Biosystems, Carlsbad, CA, USA). Actin (OsRac1, LOC_Os01g12900) was used as an endogenous control. All primers are shown in Table S2.
| Table S2. Sequences of primers for RT-qPCR. |
To construct the complementary vector ofMFS4, a fragment containing a 2 000 bp upstream sequence and complete open reading frame sequence was amplified from rice genomic DNA using the primers MFS4-com-GFP-F-EcoRI and MFS4-com- GFP-R-BamHI. The fragment was cloned into the binary vector pCAMBIA1300 at the EcoRI/BamHI site. After verification by sequence analysis, the vector was transformed into the mfs4 mutant by the Agrobacterium-mediated method. The primer sequences used are shown in Table S3.
To carry out the subcellular localization of MFS4 protein, the MFS4 coding sequence was amplified from rice genomic DNA using the primers 580-MFS4-XbalI-F and 580-MFS4-BamHI-R. The fragment was cloned into the vector pAN580 at the XbalI/BamHI site. After verification by sequence analysis, the pAN580-GFP, MTS-GFP and MFS4-GFP plasmids were transformed into rice protoplasts. After incubation at 28 º C for 12-16 h, GFP fluorescence was detected using a confocal laser scanning microscope (LSM710, Zeiss, Jena, Germany). The primers are shown in Table S3.
| Table S3. Primers used in this study. |
This study was supported by the National Natural Science Foundation of China (Grant Nos. 31971919 and 31730063), the Natural Science Foundation Project of Chongqing Science and Technology Commission, China (Grant No. cstc2020jcyj-jqX0020), National Key Program for Research and Development of China (Grant No. 2017YFD0100202), and Chongqing Graduate Research and Innovation Project Funding, China (Grant No. CYS20123).
The following materials are available in the online version of this article at http://www.sciencedirect.com/journal/rice-science; http://www.ricescience.org.
Table S1. Sequences of primers for gene mapping.
Table S2. Sequences of primers for qRT-PCR.
Table S3. Primers used in this study.
(Managing Editor: Wu Yawen)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|