Rice Science ›› 2022, Vol. 29 ›› Issue (4): 363-374.DOI: 10.1016/j.rsci.2021.11.006
收稿日期:2021-08-20
接受日期:2021-11-23
出版日期:2022-07-28
发布日期:2022-06-01
. [J]. Rice Science, 2022, 29(4): 363-374.
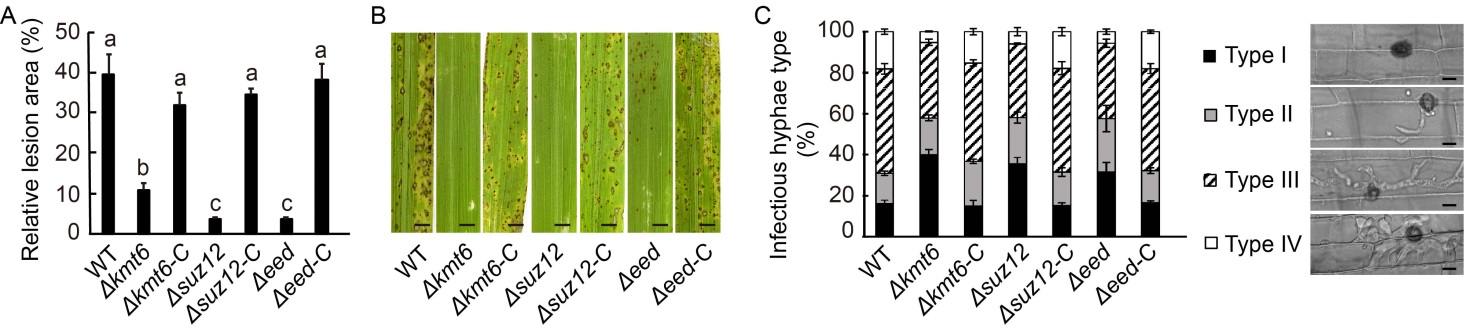
Fig. 1. Polycomb repressive complex 2 (PRC2) is required for fungal pathogenicity in Magnaporthe oryzae. A, Relative lesion area of indicated strains. The relative lesion area was quantified by an ImageJ software. Δkmt6, Δsuz12 and Δeed are deletion mutants from the wild type (WT) strain. Δkmt6-C, Δsuz12-C and Δeed-C are their complementary strains, respectively. B, Blast infection assay using 21-day-old rice seedlings (CO39). Deletion of Kmt6, Eed and Suz12 impaired the pathogenicity of M. oryzae. Scale bars, 2 mm. C, Observation and statistical analysis of invasive hypha growth in rice sheath cells at 40 h post-inoculation. Four types of invasive hyphae: no penetration (Type I), penetration with primary hyphae (Type II), with differentiated secondary invasive hyphae (Type III), and invasive hyphae spreading into neighbouring cells (Type IV), were quantified. Data represent Mean ± SD of three independent repeats, with n = 300 appressoria per analysis. Scale bars, 5 μm.
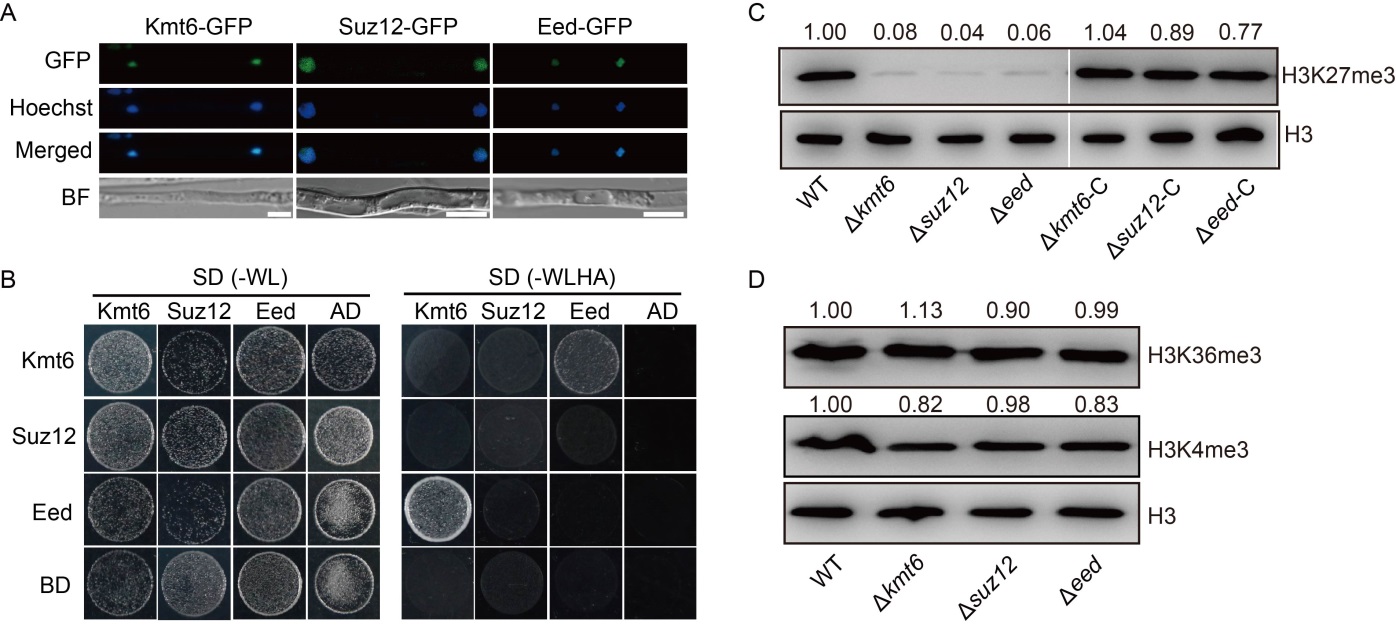
Fig. 2. Core subunits of Polycomb repressive complex 2 (PRC2) are indispensable for H3K27me3 modification in Magnaporthe oryzae. A, Confocal microscopy image for subcellular localization of PRC2 subunits fused with green fluorescent protein (GFP). The GFP signal co-localized with Hoechst (10 μg/mL) stained nuclei. BF, Blight field. Scale bars, 5 μm. B, Yeast two-hybrid assay of PRC2 components Kmt6, Eed and Suz12. The bait and prey plasmids were co-transformed into yeast strain Y2Hgold, respectively. Then, the transformants were grown on basal medium SD (-WL, without tryptophan and leucine) and selective medium SD (-WLHA, without tryptophan, leucine, histidine and adenine). The empty plasmids pGADT7 (AD) and pGBKT7 (BD) were used as controls. C, Levels of histone H3 and H3K27me3 in wild type (WT), ∆kmt6, ∆eed and ∆suz12 strains and their complementary strains (∆kmt6-C, ∆eed-C and ∆suz12-C) on total histones were measured by Western blotting. The relative intensity abundance was measured and calculated. Two repeated biological experiments were conducted with similar results. D, Levels of histone H3, H3K4me3 and H3K36me3 in WT, ∆kmt6, ∆eed and ∆suz12 strains were measured by Western blotting. The relative intensity abundances were measured and calculated. Two repeated biological experiments were conducted with similar results.
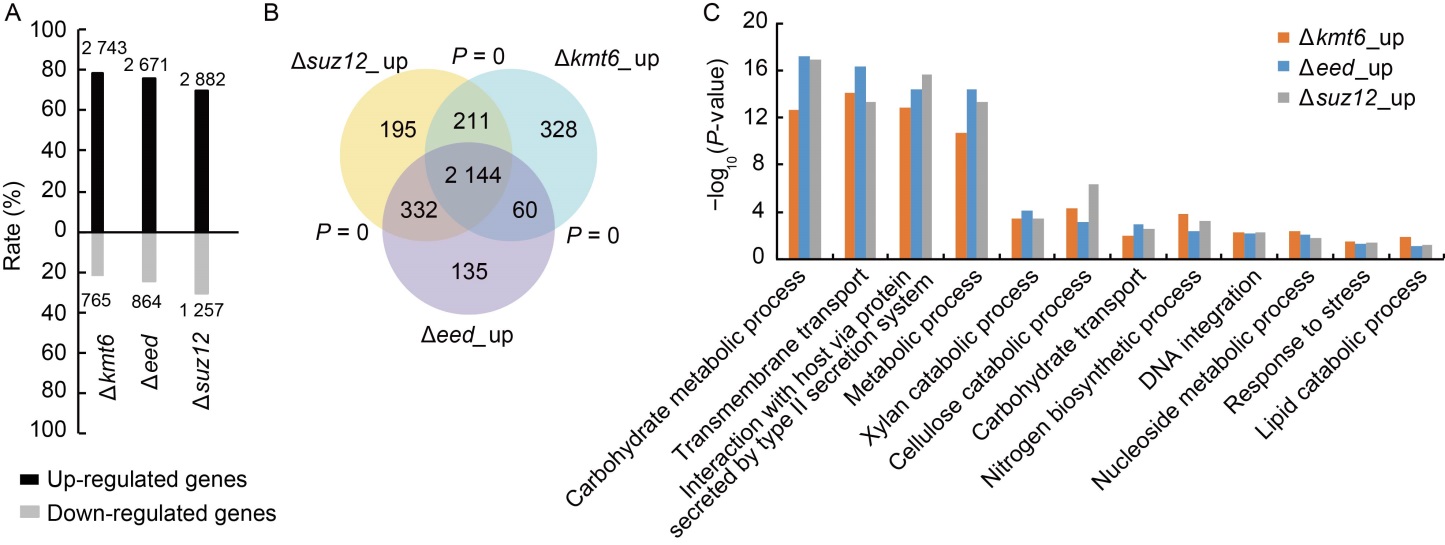
Fig. 3. Core subunits of Polycomb repressive complex 2 (PRC2) regulate genome-wide gene expression in Magnaporthe oryzae. A, Summary of up- and down-regulated genes in different PRC2 deletion mutants with RNA-seq analysis. The numbers at the top or bottom of the bars are the numbers of differential expressed genes in the mutants. The Y-axis is the percentage of up- and down-regulated genes in all the differentially expressed genes. Data were obtained from three independent biological=repeats. B, Venn diagram showing statistically significant overlaps among gene sets of ∆kmt6_up, ∆suz12_up and ∆eed_up. P value for overlapping between gene sets was obtained by the Fisher’s exact test. C, Gene Ontology (GO) analysis of ∆kmt6_up, ∆suz12_up and ∆eed_up genes.
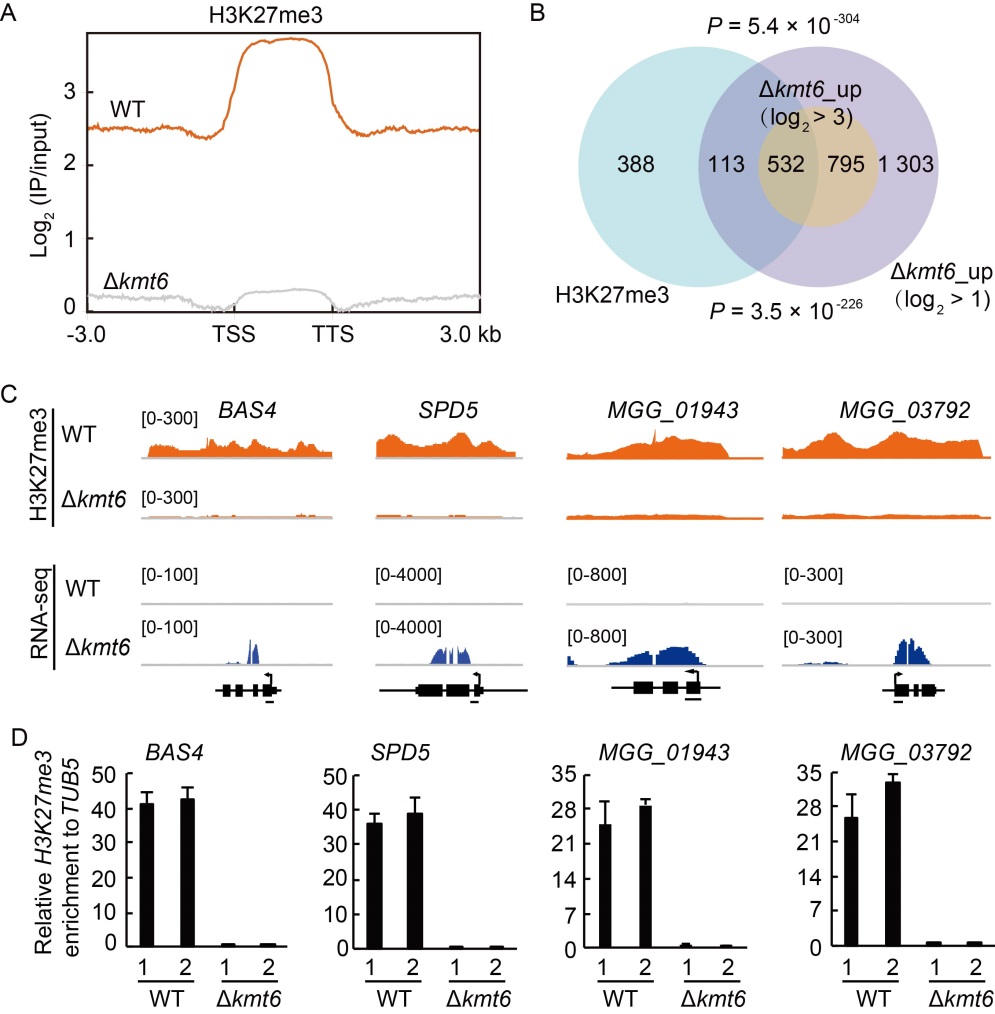
Fig. 4. H3K27me3 occupancy is highly associated with de-repression of target genes in Polycomb repressive complex 2 (PRC2) deletion mutants. A, Average H3K27me3 occupancy within 3-kb genomic regions flanking the summit of H3K27me3 peaks in wild type (WT) and ∆kmt6 strains. TSS, Transcription start site; TTS, Transcription termination site. B, Venn diagram showing statistically significant overlaps between gene sets of H3K27me3-occupied genes and ∆kmt6_up with thresholds of log2 > 1 and log2 > 3 respectively. P values with Fisher’s exact test for overlapping between gene sets were labeled. C, Genome browser views of H3K27me3 occupancy from chromatin immune- precipitation assay followed by high- throughout sequencing (ChIP-seq) and expression pattern from RNA-seq of selected genes. The number areas were reads per million (RPM). D, ChIP-qPCR verified the enrichment of H3K27me3 at the chromatin of selected genes. The examined regions were shown with black line at the bottom of gene model. The relative enrichments were calculated by relative quantitation from two biological repeats, which was standardized with an internal control TUB5, then compared with that of WT. The numbers ‘1’ and ‘2’ indicated two independent repeats. Values are Mean ± SD of three technical repeats.
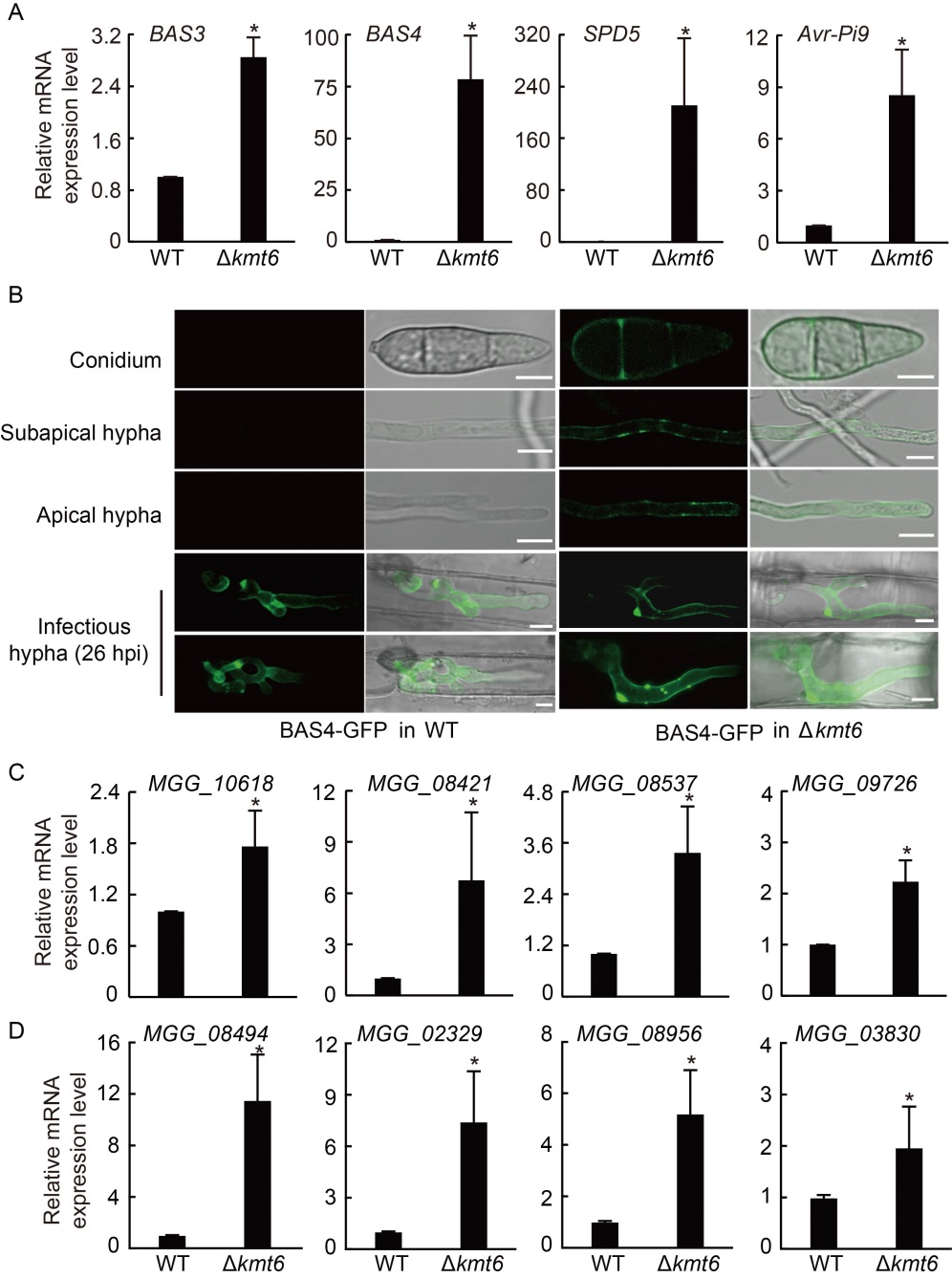
Fig. 5. Expression levels of genes which encode effector, extracellular enzyme and cytochrome P450 were up-regulated in Polycomb repressive complex 2 (PRC2) deletion mutants. A, Relative expression levels of effectors were checked in wild type (WT) and Δkmt6 strains. The strains were cultured in liquid complete medium (CM) at 28 ºC for 2 d. Tubulin was used as an internal reference. B, Confocal microscopy image of Bas4- GFP at the vegetative and in planta growth stages of WT and ∆kmt6 strains. In WT, Bas4-GFP fluorescence was only shown at the in planta growth stage, while in ∆kmt6, Bas4-GFP fluorescence can be detected in the conidia, apical hyphae, subapical hyphae and invasive hyphae. GFP, Green fluorescent protein; hpi, Hours post-inoculation. Scale bars, 5 μm. C and D, Relative expression levels of pathogenic genes that encode extracellular enzyme (C) and cytochrome P450 (D) were checked in WT and Δkmt6 strains. Tubulin was used as an internal reference. The strains were cultured in liquid CM at 28 ºC for 2 d. Values are Mean ± SD of three biological repeats. *, Significant differences at P < 0.05 between the deletion mutant and WT strains by the Student’s t-test.
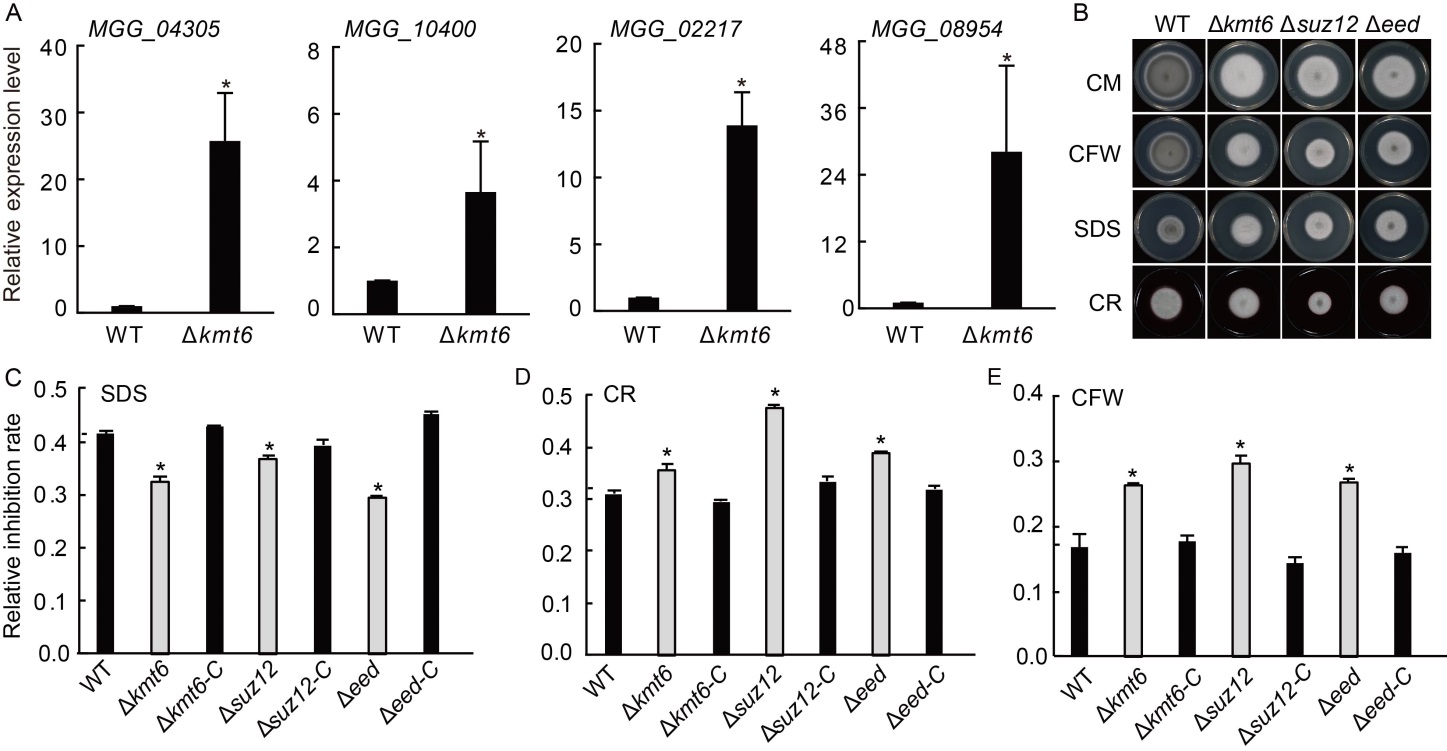
Fig. 6. Polycomb repressive complex 2 (PRC2) is required for stress response in Magnaporthe oryzae. A, Relative expression levels of cell wall related genes. Values are Mean ± SD of three biological repeats. *, Significant differences at P < 0.05 between the deletion mutant and wild type (WT) strains by the Student’s t-test. The strains were cultured in liquid CM at 28 ºC for 2 d. Tubulin was used as an internal reference. B, Radical growths of WT, ∆kmt6, ∆eed, ∆suz12 and their complementary strains (∆kmt6-C, ∆eed-C and ∆suz12-C). Colonies of indicated strains were grown on CM supplemented with different chemicals for 7 d. C?E, Statistical analysis of colony diameters of tested strains on CM supplemented with 0.005% SDS (C), 0.1 mg/mL CR (D), and 0.05 mg/mL CFW (E) for 7 d. Values are Mean ± SD of three independent repeats. *, Significant differences at P < 0.05 between the deletion mutant and WT strains by the Student’s t-test. CM, Complete medium; SDS, Sodium dodecyl sulphate; CR, Congo red; CFW, Calcofluor white.
| [1] | Blackledge N P, Rose N R, Klose R J. 2015. Targeting Polycomb systems to regulate gene expression: Modifications to a complex story. Nat Rev Mol Cell Biol, 16: 643-649. |
| [2] | Cavalli G, Heard E. 2019. Advances in epigenetics link genetics to the environment and disease. Nature, 571: 489-499. |
| [3] | Chadha S, Sharma M. 2014. Transposable elements as stress adaptive capacitors induce genomic instability in fungal pathogen Magnaporthe oryzae. PLoS One, 9: e94415. |
| [4] | Chujo T, Scott B. 2014. Histone H3K9 and H3K27 methylation regulates fungal alkaloid biosynthesis in a fungal endophyte- plant symbiosis. Mol Microbiol, 92: 413-434. |
| [5] | Connolly L R, Smith K M, Freitag M. 2013. The Fusarium graminearum histone H3 K27 methyltransferase KMT6 regulates development and expression of secondary metabolite gene clusters. PLoS Genet, 9: e1003916. |
| [6] | Ding S L, Liu W D, Iliuk A, Ribot C, Vallet J, Tao A, Wang Y, Lebrun M H, Xu J R. 2010. The Tig1 histone deacetylase complex regulates infectious growth in the rice blast fungus Magnaporthe oryzae. Plant Cell, 22: 2495-2508. |
| [7] | Dong Y H, Li Y, Zhao M M, Jing M F, Liu X Y, Liu M X, Guo X X, Zhang X, Chen Y, Liu Y F, Liu Y H, Ye W W, Zhang H F, Wang Y C, Zheng X B, Wang P, Zhang Z G. 2015. Global genome and transcriptome analyses of Magnaporthe oryzae epidemic isolate 98-06 uncover novel effectors and pathogenicity- related genes, revealing gene gain and lose dynamics in genome evolution. PLoS Pathog, 11: e1004801. |
| [8] | Dong Y H, Li Y, Qi Z Q, Zheng X B, Zhang Z G. 2016. Genome plasticity in filamentous plant pathogens contributes to the emergence of novel effectors and their cellular processes in the host. Curr Genet, 62: 47-51. |
| [9] | Dumesic P A, Homer C M, Moresco J J, Pack L R, Shanle E K, Coyle S M, Strahl B D, Fujimori D G, Yates III J R, Madhani H D. 2015. Product binding enforces the genomic specificity of a yeast Polycomb repressive complex. Cell, 160: 204-218. |
| [10] | Feng W Z, Yin Z Y, Wu H W, Liu P, Liu X Y, Liu M X, Yu R, Gao C Y, Zhang H F, Zheng X B, Wang P, Zhang Z G. 2021. Balancing of the mitotic exit network and cell wall integrity signaling governs the development and pathogenicity in Magnaporthe oryzae. PLoS Pathog, 17: e1009080. |
| [11] | Fouché S, Plissonneau C, Croll D. 2018. The birth and death of effectors in rapidly evolving filamentous pathogen genomes. Curr Opin Microbiol, 46: 34-42. |
| [12] | Galazka J M, Freitag M. 2014. Variability of chromosome structure in pathogenic fungi: Of ‘ends and odds’. Curr Opin Microbiol, 20: 19-26. |
| [13] | He M, Xu Y P, Chen J H, Luo Y, Lv Y, Su J, Kershaw M J, Li W T, Wang J, Yin J J, Zhu X B, Liu X H, Chern M, Ma B T, Wang J C, Qin P, Chen W L, Wang Y P, Wang W M, Ren Z L, Wu X J, Li P, Li S G, Peng Y L, Lin F C, Talbot N J, Chen X W. 2018. MoSnt2-dependent deacetylation of histone H3 mediates MoTor- dependent autophagy and plant infection by the rice blast fungus Magnaporthe oryzae. Autophagy, 14: 1543-1561. |
| [14] | Jamieson K, Rountree M R, Lewis Z A, Stajich J E, Selker E U. 2013. Regional control of histone H3 lysine 27 methylation in Neurospora. Proc Natl Acad Sci USA, 110: 6027-6032. |
| [15] | Kassis J A, Brown J L. 2013. Polycomb group response elements in Drosophila and vertebrates. Adv Genet, 81: 83-118. |
| [16] | Kawahara Y, Oono Y, Kanamori H, Matsumoto T, Itoh T, Minami E. 2012. Simultaneous RNA-seq analysis of a mixed transcriptome of rice and blast fungus interaction. PLoS One, 7: e49423. |
| [17] | Khang C H, Berruyer R, Giraldo M C, Kankanala P, Park S Y, Czymmek K, Kang S, Valent B. 2010. Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell movement. Plant Cell, 22: 1388-1403. |
| [18] | Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg S L. 2013. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol, 14: R36. |
| [19] | Kou Y J, Tan Y H, Ramanujam R, Naqvi N I. 2017. Structure- function analyses of the Pth11 receptor reveal an important role for CFEM motif and redox regulation in rice blast. New Phytol, 214: 330-342. |
| [20] |
Langmead B, Salzberg S L. 2012. Fast gapped-read alignment with Bowtie 2. Nat Methods, 9: 357-359.
PMID |
| [21] | Langmead B, Trapnell C, Pop M, Salzberg S L. 2009. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol, 10: R25. |
| [22] |
Lanzuolo C, Orlando V. 2012. Memories from the polycomb group proteins. Annu Rev Genet, 46: 561-589.
PMID |
| [23] | Lee J, Lee J J, Jeon J. 2019. A histone deacetylase, MoHOS2 regulates asexual development and virulence in the rice blast fungus. J Microbiol, 57: 1115-1125. |
| [24] | Li Z C, Fu X, Wang Y Z, Liu R Y, He Y H. 2018. Polycomb- mediated gene silencing by the BAH-EMF1 complex in plants. Nat Genet, 50: 1254-1261. |
| [25] | Margueron R, Reinberg D. 2011. The Polycomb complex PRC2 and its mark in life. Nature, 469: 343-349. |
| [26] | Mathioni S M, Patel N, Riddick B, Sweigard J A, Czymmek K J, Caplan J L, Kunjeti S G, Kunjeti S, Raman V, Hillman B I, Kobayashi D Y, Donofrio N M. 2013. Transcriptomics of the rice blast fungus Magnaporthe oryzae in response to the bacterial antagonist Lysobacter enzymogenes reveals candidate fungal defense response genes. PLoS One, 8: e76487. |
| [27] |
Mosquera G, Giraldo M C, Khang C H, Coughlan S, Valent B. 2009. Interaction transcriptome analysis identifies Magnaporthe oryzae BAS1-4 as biotrophy-associated secreted proteins in rice blast disease. Plant Cell, 21: 1273-1290.
PMID |
| [28] | Netea M G, Joosten L A B, Latz E, Mills K H G, Natoli G, Stunnenberg H G, O'Neill L A, Xavier R J. 2016. Trained immunity: A program of innate immune memory in health and disease. Science, 352: aaf1098. |
| [29] | Oh Y, Donofrio N, Pan H Q, Coughlan S, Brown D E, Meng S W, Mitchell T, Dean R A. 2008. Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus Magnaporthe oryzae. Genome Biol, 9: R85. |
| [30] | Pham K T M, Inoue Y, Vu B V, Nguyen H H, Nakayashiki T, Ikeda K I, Nakayashiki H. 2015. MoSET1 (histone H3K4 methyltransferase in Magnaporthe oryzae) regulates global gene expression during infection-related morphogenesis. PLoS Genet, 11: e1005385. |
| [31] | Qian B, Liu X Y, Ye Z Y, Zhou Q K, Liu P, Yin Z Y, Wang W H, Zheng X B, Zhang H F, Zhang Z G. 2021. Phosphatase- associated protein MoTip41 interacts with the phosphatase MoPpe1 to mediate crosstalk between TOR and cell wall integrity signalling during infection by the rice blast fungus Magnaporthe oryzae. Environ Microbiol, 23: 791-809. |
| [32] | Ridenour J B, Möller M, Freitag M. 2020. Polycomb repression without bristles: Facultative heterochromatin and genome stability in fungi. Genes, 11(6): 638. |
| [33] |
Robinson J T, Thorvaldsdóttir H, Winckler W, Guttman M, Lander E S, Getz G, Mesirov J P. 2011. Integrative genomics viewer. Nat Biotechnol, 29: 24-26.
PMID |
| [34] |
Sánchez-Vallet A, Fouché S, Fudal I, Hartmann F E, Soyer J L, Tellier A, Croll D. 2018. The genome biology of effector gene evolution in filamentous plant pathogens. Annu Rev Phytopathol, 56: 21-40.
PMID |
| [35] |
Schuettengruber B, Bourbon H M, di Croce L, Cavalli G. 2017. Genome regulation by polycomb and trithorax: 70 years and counting. Cell, 171: 34-57.
PMID |
| [36] | Sharpee W, Oh Y, Yi M, Franck W, Eyre A, Okagaki L H, Valent B, Dean R A. 2017. Identification and characterization of suppressors of plant cell death (SPD) effectors from Magnaporthe oryzae. Mol Plant Pathol, 18: 850-863. |
| [37] | Tao Z, Shen L S, Gu X F, Wang Y Z, Yu H, He Y H. 2017. Embryonic epigenetic reprogramming by a pioneer transcription factor in plants. Nature, 551: 124-128. |
| [38] |
Trapnell C, Williams B A, Pertea G, Mortazavi A, Kwan G, van Baren M J, Salzberg S L, Wold B J, Pachter L. 2010. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol, 28: 511-515.
PMID |
| [39] | Villalba F, Collemare J, Landraud P, Lambou K, Brozek V, Cirer B, Morin D, Bruel C, Beffa R, Lebrun M H. 2008. Improved gene targeting in Magnaporthe grisea by inactivation of MgKU80 required for non-homologous end joining. Fungal Genet Biol, 45: 68-75. |
| [40] | Wang L Y, Chen H, Li J J, Shu H D, Zhang X X, Wang Y C, Tyler B M, Dong S M. 2020. Effector gene silencing mediated by histone methylation underpins host adaptation in an oomycete plant pathogen. Nucleic Acids Res, 48: 1790-1799. |
| [41] | Wiles E T, Selker E U. 2017. H3K27 methylation: A promiscuous repressive chromatin mark. Curr Opin Genet Dev, 43: 31-37. |
| [42] | Wiles E T, McNaught K J, Kaur G, Selker J M L, Ormsby T, Aravind L, Selker E U. 2020. Evolutionarily ancient BAH-PHD protein mediates Polycomb silencing. Proc Natl Acad Sci USA, 117: 11614-11623. |
| [43] |
Xiao J, Jin R, Yu X, Shen M, Wagner J D, Pai A, Song C, Zhuang M, Klasfeld S, He C S, Santos A M, Helliwell C, Pruneda-Paz J L, Kay S A, Lin X W, Cui S J, Garcia M F, Clarenz O, Goodrich J, Zhang X Y, Austin R S, Bonasio R, Wagner D. 2017. Cis and trans determinants of epigenetic silencing by Polycomb repressive complex 2 in Arabidopsis. Nat Genet, 49: 1546-1552.
PMID |
| [44] | Yang Z L, Qian S M, Scheid R N, Lu L, Chen X S, Liu R, Du X, Lv X C, Boersma M D, Scalf M, Smith L M, Denu J M, Du J M, Zhong X H. 2018. EBS is a bivalent histone reader that regulates floral phase transition in Arabidopsis. Nat Genet, 50: 1247-1253. |
| [45] | Zhang S L, Liang M L, Naqvi N I, Lin C X, Qian W Q, Zhang L H, Deng Y Z. 2017. Phototrophy and starvation-based induction of autophagy upon removal of Gcn5-catalyzed acetylation of Atg7 in Magnaporthe oryzae. Autophagy, 13: 1318-1330. |
| [46] | Zhang W, Huang J, Cook D E. 2021. Histone modification dynamics at H3K27 are associated with altered transcription of in planta induced genes in Magnaporthe oryzae. PLoS Genet, 17: e1009376. |
| [47] | Zhang Y, Liu T, Meyer C A, Eeckhoute J, Johnson D S, Bernstein B E, Nusbaum C, Myers R M, Brown M, Li W, Liu X S. 2008. Model-based analysis of ChIP-Seq (MACS). Genome Biol, 9: R137. |
| No related articles found! |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||