Rice Research Institute, Key Laboratory of Application and Safety Control of Genetically Modified Crops, Academy of Agricultural Sciences, Southwest University, Chongqing 400715, China
版权声明:
2021 Editorial Office of Rice Science China National Rice Research Institute
作者简介:
#These authors contributed equally to this work
展开
Abstract
Inflorescence structure of rice, including the number and length of branches, and the density of the spikelet, can greatly affect the number of grains per panicle, which is one of the key factors in yield compositions. Here we identified five allelic mutants sb1-1/2/3/4/5 that related to branch development of rice. In these mutants, the branch meristem fate was prolonged sharply, resulting in delay of transition from branches to spikelets, and then increased the numbers of branches and spikelets per panicle. SB1 encodes a nuclear RING-like domain protein of SHI/LRP/SRS family and strongly expressed in branch meristems. The results of protein interaction and chromatin immunoprecipitation further suggested that SB1 directly repressed the expression of DEP1, TAW1, MOC1 and IPA1 by interacting with a co-repressor complex to affect acetylation level of histone H3 on target regions. Thus, we proposed that SB1 is a transcription repressor of branch meristem activity by widely and negatively regulating a series of genes that maintain branch meristem fate.
Keywords:branch development
;
grain number per panicle
;
rice
;
RING-like zinc-finger
;
transcription repressor
;
panicle arthitecture
XiaoqinZeng, HuiZhuang, QinglanCheng, JunTang, FayuYang, MingjiangHuang, ZiyiWang, ZhongchengLi, HonghuiZhu, RuiChen, GuanghuaHe, YunfengLi. SB1 Encoding RING-Like Zinc-Finger Protein Regulates Branch Development as a Transcription Repressor[J]. Rice Science, 2021, 28(3): 243-256 https://doi.org/10.1016/j.rsci.2021.04.004
In grass crops, grain number per panicle (GNP) is one of the most important yield components (Wang and Li, 2011). Panicle architecture consisting of the number of branches, the length of branches, and grain density on branches, determines the GNP (Li et al, 1998; Teng et al, 2002). In the past two decades, dozens of genes related to panicle architecture have been characterized in rice, some of which play crucial roles in determining GNP and then yield.
In rice, primary branches (PBs), secondary branches (SBs) and lateral spikelets (LSs) are all originated from axillary meristem, which compose the panicle with main axis and topic spikelet (TS) originating from apical meristem. Therefore, the genes participating in the initiation, formation and maintaining of axillary meristem in panicle are first regulators for panicle architecture and GNP, such as LAX PANICLE1 (LAX1), LAX2 and MONOCULM1 (MOC1) in rice. They all encode transcription factor, and the loss-of-function mutants of them all display sharp decreasing of branch and spikelet number (Li et al, 2003; Skirpan et al, 2008; Tabuchi et al, 2011). It is proved that LAX1 can interact with LAX2 and MOC1 respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development.
The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017).
Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016).
Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018).
To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice.
RESULTS
Characterization of sb1 mutant phenotype
We identified five allelic recessive mutants named sb1-1/2/3/4/5. Compared with the wild type plants, the mutants showed no significant difference in plant height or tiller number but the panicles of sb1 generated more branches. A mature wild type rice panicle was composed of a main axis, branches [PBs, SBs, barely tertiary branches (TBs)] and spikelets (TS and LS) produced on the branches (Fig. 1-A and -B). In contrast, although there was no significant difference in the numbers of PBs between sb1-1/2/3/4/5 and the wild type (Fig. 1-C), the numbers of SBs, TBs and spikelets per panicle of the five allelic mutants were significantly increased (Fig. 1-D to -F). Among them, the numbers of SBs of sb1-1/2/3/4/5 reached 92 (+104.4%), 59 (+31.1%), 79 (+75.5%), 64 (+42.2%) and 68 (+51.1%), respectively, whereas it was only 45 in the wild type (Fig. 1-D). Particularly, the wild type panicle usually produced few TBs, while the sb1-1/3/5 produced 84, 34 and 13 TBs, respectively (Fig. 1-E). The numbers of spikelets per panicle were increased remarkably in the sb1 mutants. There were the highest number of spikelets per panicle in sb1-1 with an average of 473 (+125.2%), and the lowest number in sb1-2 with an average of 291 (+38.6%), whereas the average number of spikelets in the wild type was only 210 (Fig. 1-F). However, the increase of spikelet numbers in the sb1 mutants did not increase grain number, and the sb1-1/2/3/4/5 mutants generated only about 82, 175, 130, 190 and 176 grains per panicle, respectively. Some of them were significantly lower than the average number of grains in the wild type (190 grains per panicle) (Fig. 1-G). It was because the seed-setting rates of sb1-1/2/3/4/5 were only 17.7%, 64.8%, 33.6%, 70.0% and 56.4%, which were significantly lower than 90.2% in the wild type (Fig. 1-H).
Fig. 1. Phenotypes of panicles in wild type (WT) and sb1-1/2/3/4/5 mutants. A, Panicles of the WT and sb1-1/2/3/4/5 mutants with spikelets on the branches. Bars are 5 cm. B, Panicles of the WT and sb1-1/2/3/4/5 mutants without spikelets on the branches. Bars are 5 cm. C‒H, Numbers of primary branches per panicle (C), numbers of secondary branches per panicle (D), numbers of tertiary branches per panicle (E), numbers of spikelets per panicle (F), numbers of grains per panicle (G) and seed-setting rates (H) of WT and sb1-1/2/3/4/5. The wild type plants and sb1-1/2/3/4/5 mutants were grown in standard paddy field under conventional cultivation conditions. Data are Mean ± SD (n = 10). The Student’s t-test was used to generate the P values. **, P < 0.01.
In addition, we observed the development of branches and spikelets in young panicles using a scanning electron microscopy. As expected, more branch primordia were observed in the sb1-1 panicles (about 0.5 cm in length), while the branch primordia rarely appeared at the similar stage of the wild type panicles (Fig. S1-A to -D). Next, we detected the expression of genes that involving in the BM activity. The results showed that the expression levels of LAX1, RCN1, OSH1 and MOC3 were significantly upregulated in the sb1-1 panicles (Fig. S1-E). Therefore, our data suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets.
Fig. S1. scanning electron microscopy analysis of young panicles of sb1-1 mutant and wild type. A, Wild type panicles. B, Magnified figure of A. C, sb1-1 panicles. D, Magnified figure of C. E, Expression of LAX1, RCN1, OSH1 and MOC3. bm, branch meristem; sm, spikelet meristem. Bars are 500μm. Data are Mean ± SD (n = 3). The Student’s t-test was used to generate the P values. *, P < 0.05, **, P < 0.01.
Fig. S3. Protein sequence analysis of SHI/LRP/SRS protein family in rice, maize and Arabidopsis thaliana.
Map-based cloning of SB1
All F1 plants of the cross 56S × sb1 showed normal phenotype. In the F2 population, 2 874 plants were obtained in total, among which 2 198 plants showed normal phenotype and 693 plants with mutant phenotype. Genetic analysis indicated that the numbers of wild type and mutant individuals fit the expected 3:1 ratio (χ2 = 1.73 < χ20.05 = 3.84), which meant the sb1 phenotype was controlled by a single recessive gene.
To identify the SB1 gene, the 693 mutated individuals were used as mapping population. Linkage analysis revealed that the SB1 locus was associated with the simple sequence repeat (SSR) markers SSR1 and SSR2 on the long arm of chromosome 9. Subsequently, the candidate region was further delimited to a 122-kb interval between two single nucleotide polymorphism (SNP) primers SNP-1 and SNP-2, where 13 annotated genes were predicted (Fig. 2-A). Sequencing analysis revealed that a G-T substitution in the last base of the first exon of LOC_Os09g36160 occurred in the sb1-1 mutant. The mutation caused the 183th codon changed into termination codon and led to premature translation termination. Sequencing analysis also revealed that other four mutants had mutational sites in the candidate gene LOC_Os09g36160. A 29-bp fragment deletion since the 166th base and a 4-bp deletion since the 298th base occurred in sb1-2 and sb1-3, respectively, which both led to frameshift mutation. sb1-4 had a T-A substitution on the 661th base of the second exon and led to Tyr-Asn substitution of the 221th amino acid. sb1-5 had a 51-bp fragment deletion since the 664th base and led to 17 amino acids deletion in the SB1 protein (Fig. 2-B). To verify whether LOC_Os09g36160 was responsible for the sb1 phenotype, a 4091-bp SB1-GFP (green fluorescent protein) fusion expression vector which contained 3070-bp upstream sequence and 1021-bp SB1 open reading frame (ORF) sequence without termination codon was transformed into sb1-1 (Fig. 2-C). Totally, 17 transgenic plants were obtained, and we then analyzed the coding sequence of LOC_Os09g36160 and examined the GFP signal to identify the positive plants. Among them, 11 plants showing double peaks at the mutant site also showed obvious GFP fluorescent signal, which revealed that the combination plasmid was successfully transformed into these 11 plants. All these 11 transgenic plants displayed normal panicles with similar numbers of branches and spikelets per panicle with those in the wild type (Fig. 2-D). These results finally confirmed that LOC_Os09g36160 gene was indeed the SB1 gene.
Fig. 2. Map-based cloning of SB1. A, Fine mapping of SB1 on chromosome 9. B, Structure of SB1 and mutation sites of sb1-1/2/3/4/5. C, Structure of the SB1-GFP fusion complementary vector. D, Phenotypes of the wild type, sb1-1 and sb1-1-GFP transgenic plant. Bars are 5 cm. ORF, Open reading frame; SB1-comF and SB1-comR, Cloning primers of SB1 for complementary test.
SB1 gene encodes a nuclear-located SHI/LRP/SRS family protein
SB1 encodes a RING-like zinc finger transcription factor that belongs to SHI/LRP/SRS family. In Arabidopsis, the SHI family contains 10 members: SHORT INTERNODES (SHI), STYLISH1 and 2 (STY1 and STY2), SHI-RELATED SEQUENCE3 (SRS3) to SRS8, and LATERAL ROOT PRIMORDIUM (LRP) (Fridborg et al, 2001). Phylogenetic analysis revealed that there were five SHI/LRP/SRS protein family members in rice. SB1 had a poor homology with all SHI/LRP/SRS proteins in Arabidopsis and had a close evolutionary relationship with ZmLRL4 and ZmLRL5 in maize.
The LOC_Os01g724901 and LOC_Os05g320701 had high homology and close branching relationship with LRP, which was widely studied in Arabidopsis and maize (Fig. 3-A). Previous studies have shown that the sequences of the SHI/LRP/SRS family proteins were highly diverse, with highly conserved regions: a C3HC3H RING-like domain and an IGGH domain (Fridborg et al, 2001; Kuusk et al, 2002, 2010). Then, the protein motif prediction program MEME and protein sequence alignment program DNAMAN were used to analyze the full-length amino acid sequences of SHI/LRP/SRS family proteins in rice, maize and Arabidopsis. The result showed that there were three highly conservative domain/motifs: a RING-like structure domain, an IGGH domain and an FRCVR domain in the SHI/LRP/SRS family proteins. In previous results, the FRCVR domain was not identified separately (Figs. 3-B, S2 and S3).
Fig. 3. SB1 encodes a RING-like zinc finger protein. A, Phylogenetic tree analysis. The phylogenetic tree was constructed using the neighbor- joining method based on the Jone-Taylor- Thornton matrix-based model, and bootstrap support values calculated from 1 000 replicates are given at the branch nodes. At, Arabidopisis thaliana; Os, Oryza sativa; Zm, Zea mays. B, Location of conserved domains of the genes in the phylogeny tree.
To confirm the subcellular localization of SB1 protein, the fluorescence signal of the SB1-GFP fusion protein was observed directly in the SB1P:SB1:GFP complementary transgenic plants using a confocal laser scanning microscope. The GFP signal of SB1-GFP fusion protein was localized in the nucleus (Fig. S4). These results together demonstrated that SB1 was a nuclear-located SHI/LRP/SRS family transcription factor.
Fig. S4. Subcellular location of SB1 by observing the GFP signal in SB1P:SB1: GFP transgenic plants. A, GFP signals in cells of branch meristem. B, DAPI signal in nucleus. C, Merged fluorescence signal of GFP and DAPI. Bars are 50 μm.
Spatiotemporal expression pattern of SB1
Firstly, the expression of SB1 gene in different rice tissues was detected by qPCR. The results revealed that SB1 gene was strongly expressed in the young panicle, while it was weakly expressed in the shoot, tiller bud, leaf and sheath, and almost not expressed in the root (Fig. 4-A). Furthermore, we analyzed the signal of GFP in detail using young panicles of the SB1P:SB1:GFP complementary transgenic lines (Fig. 4-B to -M). Strong GFP signals were first observed in the primary branch primordia at the inflorescence stage In2 (Fig. 4-B and -C), and then the GFP signal was enhanced as the number of primary branch primordia increased during the stage In3 (Fig. 4-D and -E). During the stages In4‒5, when the PBs continuedly elongated and generated secondary branch primordia, strong GFP signal was observed in the second branch primordia as expected (Fig. 4-F to -I). With the growth of primary and secondary branches, the GFP expression was still observed in the elongated SBs and the potential ancestral cells of SM (Fig. 4-J to -M). These results indicated that during panicle development, SB1 expression was always strongly detected in the branch primordia, which was well corresponded with the mutant phenotype of increased branch number.
Fig. 4. Expression pattern of SB1. A, qPCR analysis of SB1 in root, shoot, leaf, sheath, tiller bud and panicles of different length. Panicles 1, 2 and 3 refer to panicles with the lengths of < 0.5, 0.5‒1.0 and 1.1‒2.0 cm, respectively. OsActin was used as a reference gene. Data are Mean ± SD (n = 3). B‒M, GFP signal indicating SB1 expression in complementary transgenic plants harboring the construct SB1P: SB1:GFP. B and C, Panicles at the stage In2; D and E, Panicles at the stage In3; F and G, Panicles at the stage In4; H and I were the magnification figure of a PBM in F and G; J and K, Panicles at the stage In5; L and M were the magnification figure of a PBM in J and K. PBM, Primary branch meristem; SBM, Secondary branch meristem; SM, Spikelet meristem. In2, 3, 4 and 5 refer to the four inflorescence stages of rice during which the branch meristems formed. Bars are 100 μm.
SB1 interacts with transcriptional corepressor complex
To illuminate the function of SB1, we carried out a yeast two- hybrid (Y2H) assay to find the interacting proteins of SB1. Firstly, the full-length SB1-BD plasmid was transformed into Y2HGold yeast cells. The yeast cells that harboured pGBKT7-FZP (positive control) can grow on both SD/-Trp and SD/-Trp-His-Ade media, whereas the yeast cells that harboured pGBKT7-SB1 and the empty pGBKT7 can only grow on the SD/-Trp medium but not on the SD/-Trp-His-Ade medium, indicating that SB1 protein showed no auto-activation ability (Fig. 5-A). Then, pGBKT7-SB1 was used as a bait to screen its partners in a cDNA library from rice young panicles. According to the screening results, we found two proteins: Os06g0126000 (OsSEUSS-like 1, OsSEU1) and Os04g0510200 (OsLEUNIG-like 1, OsLUG1) (Table S1), which are annotated as transcriptional corepressors and are homologous of SEU and LUG in Arabidopsis (http://rice.plantbiology. msu.edu/). Previous research had shown that the corepressor complex SEU-LUG plays an important role in plant development (Conner and Liu, 2000; Franks et al, 2002; Sridhar et al, 2004; Liu and Karmarkar, 2008; Sitaraman et al, 2008). Then, a Y2H assay was performed to test the interaction of SB1 with OsSEU1 and OsLUG1. As expected, after about 3 d, the yeast that co-expressed SB1 and OsLUG1 grew on the SD/-Trp-Leu-His-Ade selective medium, and the yeast that co-expressed SB1 and OsLUG1 grew after about 5 d. The results revealed that SB1 can interact with OsLUG1 and OsSEU1, and the interaction between SB1 and OsSEU1 was weaker than that of SB1 and OsLUG1 (Fig. 5-B). Next, a bimolecular fluorescent complimentary (BiFC) assay was conducted to further detect whether the SB1 can interact with SEU and LEU in leaf epidermal cells of Nicotiana benthamiana. Yellow fluorescent protein (YFP) signal was found in the nuclear of epidermal cells of tobacco leaves when co-expressing the SB1-YFPC fused protein with OsSEU1-YFPN and OsLUG1-YFPN fused proteins, respectively (Fig. 5-C). The results revealed that SB1 was capable of interaction with OsSEU1 and OsLUG1 in plant cells, and then acted as a transcriptional repressor in regulating the downstream genes.
Fig. 5. Interaction of SB1 with corepressors of OsSEU1 and OsLUG1. A, Autoactivation analysis of SB1 protein. PGBKT7-FZP was used as a positive control and PGBKT7-empty as a negative control. B, SB1 interacted with OsSEU1 and OsLUG1 in yeast cells by a yeast two- hybrid assay. C, SB1 interacted with OsSEU1 and OsLUG1 in epidermal cells of Nicotiana benthamiana by a bimolecular fluorescent complimentary (BiFC) assay. YFP, Yellow fluorescent protein; ChlA, Chlorophyll A; DAPI, 4’,6-diamidino-2-phenylindole. Bars are 20 μm.
Table S1.
Table S1. Interacting proteins of SB1 identified by yeast two-hybrid screening.
No.
ID
Description
1
Os09g0552800
No apical meristem protein, putative
2
Os12g0580700
Zinc finger, C3HC4 type domain containing protein
3
Os05g0103500
CHCH domain containing protein
4
Os03g0685600
WD40 repeat-like domain containing protein
5
Os01g0231000
OsIAA3 - Auxin-responsive Aux/IAA gene family member
6
Os02g0757100
Cell cycle gene
7
Os09g0567000
DUF1618 domain containing protein
8
Os02g0170500
Nuclear transcription factor Y subunit C
9
Os02g0261100
Licitor-responsive ubiquitin-conjugating enzyme
10
Os04g0510200
Transcriptional corepressor LEUNIG
11
Os06g0160400
Heading and grain weight regulat gene
12
Os02g0203000
bZIP transcription factor domain containing protein
SB1 directly and negatively regulates expression of DEP1, TAW1, MOC1 and IPA1
To clarify the mechanism of SB1 regulating branch development, chromatin immunoprecipitation-sequence (ChIP-seq) was performed to screen the direct down- stream target genes of SB1 using the young panicles of SB1P:SB1: GFP complementary transgenic lines with a GFP-antibody. A total of 3 535 significant binding peaks which covering 2 323 genes were screened by data analysis (fold enrichment > 2 and q-value < 0.001). Among these 2 323 genes, several genes such as DEP1, TAW1, MOC1 and IPA1 that were known to regulate rice branch development showed obvious potential binding peaks with SB1. There were three binding peaks in the promoter of DEP1 and one peak in the coding sequence of DEP1, one peak in the 3′-untranslated regions of TAW1, two peaks in the coding sequence of MOC1 and one peak in the promoter of IPA1 (Fig. 6-A). Next, we detected the expression levels of these four genes in the young panicles of the wild type and sb1 mutants. qPCR results revealed that the expression levels of DEP1, TAW1 and MOC1 were significantly increased among different lengths of panicles in the sb1-1 and sb1-2 mutants. The expression level of IPA1 was significantly increased in panicles (< 0.5 cm) while slightly increased in other panicles (Fig. 6-B). To further confirm whether SB1 directly regulated the expression of these genes, ChIP-qPCR was used to examine the binding between SB1 with these four genes. Chromatin isolated from young panicles of SB1P:SB1:GFP transgenic lines was immunoprecipitated with a GFP antibody and then subjected to qPCR analysis. The results verified that SB1 can directly bind with DEP1 (P2 site), TAW1 (P2 site), MOC1 (P2 site) and IPA1 (P1 site) (Fig. 6-C). Therefore, our data indicated that SB1 directly and negatively regulated the expression of DEP1, TAW1, MOC1 and IPA1 genes.
Fig. 6. SB1 directly repressed expression of DEP1, TAW1, MOC1 and IPA1. A, Binding peaks of SB1 with DEP1, TAW1, MOC1 and IPA1 by chromatin immunoprecipitation-sequence (ChIP-seq) using an anti-GFP antibody. B, Relative expression of DEP1, TAW1, MOC1 and IPA1 in the wild type (WT) and sb1 panicles, respectively. C, ChIP-qPCR for P1 and P2 sites of DEP1, TAW1, MOC1 and IPA1 with anti-GFP antibody, respectively. ChIP enrichment compared with the input sample was tested by qPCR. D, ChIP-qPCR for several sites of DEP1, TAW1, MOC1 and IPA1 with a H3K9Ac antibody, respectively. ChIP enrichment compared with the IgG sample was tested by qPCR. Pro, Promoter; P1 to P6 represent primers for ChIP-qPCR. Error bars indicated SD of three repeats. The Student’s t-test was used to generate the P values. *, P < 0.05; **, P < 0.01.
Furthermore, it is reported that the LUG and SEU complex can interact with histone deacetylases to mediate the acetylation level of histone protein in the targeted region (Gonzalez et al, 2007; Liu and Karmarkar, 2008). We further detected the acetylation levels of histone protein H3 in the promoter and coding sequence region of DEP1, TAW1, MOC1 and IPA1 by ChIP-qPCR with an anti-H3K9Ac antibody. The result indicated that the acetylation levels of H3K9 in DEP1 (P1, P2 and P3 sites), TAW1 (P2 and P3 sites), MOC1 (P1, P4 and P5 sites) and IPA1 (P1 and P2 sites) were significantly increased in the sb1 mutant compared with those in the wild type (Fig. 6-D). These results suggested that SB1 regulated the transcription of DEP1, TAW1, MOC1 and IPA1 genes by the modification of histone acetylation.
DISCUSSION
SB1 is an important factor regulating GNP in rice
In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010).
In this study, all the five mutants sb1-1/2/3/4/5 were manifested as extremely significant increase in the numbers of SBs and TBs (which barely appeared in the wild type plants), and finally showed an increase in the number of spikelets per panicle. Among them, the average ‘number of spikelets per panicle’ of sb1-1 reached nearly twice as much as the wild type. However, it was regretful that the number of grains per panicle in the sb1 mutant was obviously decreased because of the sharply decreased seed-setting rate. That meant the increasing numbers of spikelets in the sb1 mutant was meaningless unless handling the defect of seed-setting rate. In fact, it was reported that the advantageous traits (such as the increased number of branches) and disadvantageous factors (such as abnormal seed-setting rate and defect floral organs) for grain yield were concurrent in some mutants of panicle-related genes. For example, in the mutants of OsMADS34/PAP2, although the number of SBs increases sharply, the development of spikelet is affected and the seed-setting rate decreases (Gao et al, 2010; Kobayashi et al, 2010). Therefore, while the excellent natural variations in the genes such as Gn1a, TAW1, DEP1 and IPA1 had been used to largely benefit grain yield in rice, rebuilding and utilization of the genes pros and cons coexisting like SB1 and OsMADS34/PAP2 by the molecular design might be a future direction with quite development potentiality for the panicle architecture and grain yield in rice breeding.
Furthermore, the phenotypes of sb1-1/2/3/4/5 showed difference to some degree, such as the numbers of branches, spikelets and grains per panicle. Among the five mutants, sb1-1 and sb1-3 generated more branches and spikelets on panicle but less grains. Premature translation termination in sb1-1 caused the missing of the FRCVR domain and IGGH domain of SB1, and the frameshift mutation in sb1-3 caused severe destruction of the SB1 structure, which may lead to the loss of SB1 protein function. By contrast, sb1-4 and sb1-5 only had single amino acid substitution and 17-amino acid deletion in the IGGH domain, respectively, and the phenotype was significantly weaker than sb1-1/3. Interestingly, the deletion of 29-bp on the first exon in sb1-2 led to the frameshift mutation, but the new protein was highly similar to SB1, and its phenotype was similar to sb1-4/5. That is, the integrity of SB1 protein is necessary for the normal development of rice panicle, and the severity of domain deletion or destruction is positively correlated with the mutant phenotype.
SB1 might accelerate transition from BM to SM as a broad-spectrum repressor
In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets.
SB1 gene encodes a SHI/STY/SRS family protein and is allelic to OsSHI1. OsSHI1/SB1 gene regulates branch development in rice by acting as a repressive partner of IPA1 protein, a transcription activator of DEP1, to down-regulate the expression of DEP1 (Duan et al, 2019). In shi1, the mutant of complete- deletion of OsSHI1 gene, the numbers of secondary branches and spikelets increase sharply, and the DEP1 gene is also up-regulated significantly, similar with the sb1 mutants. In the present study, we showed a more precise mechanism of OsSHI1/SB1 regulating branch development. Firstly, the data of ChIP-seq and ChIP-qPCR showed SB1 can directly bind to the promoters of multiple panicle-related genes including TAW1, MOC1 and IPA1, besides DEP1. Secondly, we further revealed that SB1 can interact with two proteins of the SEU-LUG co-repressing complex, OsSEU1 and OsLUG1. SEU-LUG is known as a plant Gro/Tup1 co-repressor complex and responsible for transcriptional repressing in multiple development events. In Arabidopsis, the LUG-SEU complex with HDA19 can repress the AGMOUS (AG) expression in outer two whorls of floral organs via removing the acetylation mark from the AG locus (Conner and Liu, 2000; Liu and Karmarkar, 2008; Guo et al, 2015). Next, we also found the H3K9Ac level was increased in multiple sites of the ChIP-target genes. Therefore, our results together supported that SB1 can interact with the co-repressing complex OsSEU1-OsLUG1 and then directly repressed the expression of IPA1, DEP1, TAW1 and MOC1 through affecting the histone H3 acetylation level on target regions. In particular, the result supported that SB1 acted as a broad- spectrum transcriptional repressor on multiple targets in regulating panicle architecture in rice. Then, we speculated, it was the releasing of multiple branch development genes including IPA1, DEP1, TAW1 and MOC1 that eventually resulted in extremely prolonging BM fate and then dramatically increasing the numbers of spikelets in the sb1 and shi1 mutants (Fig. 7).
Fig. 7. Model of SB1 gene regulating rice panicle architecture. In the wild type plants, SB1 interacts with the corepressors OsSEU1 and OsLUG1 to form the repression complex and repress DEP1, TAW1, MOC1 and IPA1 by downregulating the acetylation levels of histone on chromosome. In the sb1-1/2/3/4/5 mutants, the repression complex cannot work, resulting in the increased acetylation levels and the expression levels of DEP1, TAW1, MOC1 and IPA1, which enables the formation of the increased numbers of branches and spikelets. HDACs, Histone deacetylases; BM, Branch meristem; PB, Primary branch; SB, Secondary branch; TB, Tertiary branch.
METHODS
Rice materials
The sb1-1/2/3/4/5 mutants were derived from an ethyl methane sulfonate (EMS) mutagenesis of an indica rice cultivar XIDA1B. The sb1 mutant was crossed with a sterile line 56S (indica) to yield the F1 population and subsequently generate the F2 mapping population. All rice materials were grown in experimental fields at the Rice Research Institute of Southwest University, China under natural conditions.
Phenotype characterization of sb1 mutant
During the maturity period, the complete panicles of the sb1-1/2/3/4/5 and wild type plants were photographed using a single lens reflex camera (Canon, Tokyo, Japan). Ten main panicles of the sb1-1/2/3/4/5 and wild type plants were then selected respectively to investigate phenotypical data, including the numbers of primary/secondary/tertiary branches, the number of spikelets/grains per panicle and the seed-setting rate. The development progress of young panicles (sb1 and wild type plants) was examined using a SU3500 scanning electron microscope (Hitachi, Tokyo, Japan) at -20 ºC under a low- vacuum environment.
Map-based cloning of SB1
The 693 F2 plants that exhibited a mutant phenotype were selected as a mapping population. Locus polymorphic SSR markers that developed by our laboratory were employed for gene mapping of SB1. Candidate genes were predicted using the Rice Genome Brower website (http://rice.plantbiology.msu.edu) and cDNAs of the genes in the fine-mapped region were amplified from both the wild type and the sb1 mutant, and the PCR products were confirmed by sequencing in the TSINGKE biological technology company (Chongqing, China). The primers used in the mapping and candidate gene analysis were listed in Table S2.
To construct a complementary plasmid, a 4091-bp genomic fragment that contained the 3070-bp upstream sequence and 1021-bp SB1 coding sequence coupled with GFP reporter gene coding sequence was amplified using the primers SB1-comF and SB1-comR (Table S2). The fragment was inserted into a binary vector pCAMBIA1300 using the pEASY®-Uni Seamless Cloning and Assembly Kit (TRANSGENE, Beijing, China) to construct the pCAMBIA1300-SB1-GFP vector. The recombinant plasmids were transformed into the sb1 mutant using the Agrobacterium tumefaciens-mediated transformation method as described previously (Xiao et al, 2009).
RNA isolation and qPCR analysis
Total RNAs from the root, shoot, leaf, sheath, tiller bud and panicles of the wild type plant were isolated using the RNAprep Pure Plant Kit (Tiangen, Beijing, China). The first-strand complementary cDNA was synthesized from 2 µg total RNA based on the PrimeScript® Reagent Kit with gDNA Eraser Kit (Takara, Dalian, China). qPCR was performed using the SYBR® Premix Ex Taq™ II Kit (Takara, Dalian, China) in an ABI 7500 Real-time PCR System (ABI, Carlsbad, USA). Relative gene expression was quantitated based on three biological replicates and OsActin (OsRac1, LOC_Os01g12900) was used as an endogenous control. The sequences of primers used in qPCR were listed in Table S2.
Subcellular localization
We confirmed the subcellular localization of SB1 according to the SB1-GFP fluorescence in the positive complementary transgenic plants. At the booting stage, single branch or spikelet of young panicles were transferred to a glass slide, then examined the fluorescence signal under an Olympus FluoView 1000-Confocal laser scanning microscope (ZEISS, Jena, Germany).
Protein sequence and phylogenetic analysis
The SB1-related protein sequences were downloaded from GenBank (http://www.ncbi.nlm.nih.gov/genbank/) using the full-length amino acid sequence of SB1 as a query. A phylogenetic tree was constructed using the maximum likelihood method based on the Jones-Taylor-Thornton matrix-based model with the lowest Bayesian information criterion scores by MEGA 5.0 (Tamura et al, 2011). Statistical support for the tree topology was assessed by means of a bootstrap analysis with 500 replicates. Protein motifs were predicted using the motif-based sequence analysis tool MEME (http://alternate.meme-suite.org/).
Yeast two-hybrid (Y2H) assay
The full-length SB1 coding region was cloned into a pGBKT7 vector at the EcoRI and BamHI restriction sites. Then the pGBKT7-SB1 construct was transformed into the yeast strain Y2HGold. Transformants were selected on SD/-Trp and SD/-Ade/-His/-Trp media to test the auto-activation of SB1. FZP protein having strong auto-activation was used as a positive control and the empty pGBKT7 vector as a negative control. A young inflorescences cDNA library constructed by Shanghai OE Biotechnology Co., Ltd in China was used to perform the Y2H screening and positive clones were identified by sequencing. The full-lengths of OsSEU1 and OsLUG1 coding regions were cloned into the pGADT7 vector at the EcoRI and BamHI restriction sites, respectively. Then the pGADT7-OsSEU1 and pGADT7-OsLUG1 were used as preys and separately transformed into the yeast strain Y2HGold together with the bait construct pGBKT7-SB1. The selective medium SD-Leu/-Trp/-His/-Ade was used to test the interaction between bait (SB1) and preys (OsSEU1 and OsLUG1). The sequences of primers used in Y2H assay were listed in Table S2.
Bimolecular fluorescent complimentary (BiFC)
To conduct BiFC assay, the coding sequences of SB1, OsLUG1 and OsSEU1 were amplified by gene-specific primers (Table S2) and then ligated into BiFC vectors, including pSCYNE (SCN, modified) and pSCYCE (SCC, modified). The recombinant vector pairs were co-transformed into N. benthamiana leaves. The fluorescence signals were captured under a confocal laser scanning microscope (LSM 800, Zeiss, Germany).
ChIP-Seq and ChIP-qPCR
The young panicles (< 3 cm) from the SB1P:SB1:GFP complementary transgenic plants were collected to do the ChIP-seq using a GFP-antibody. The sequencing and data analysis were carried out by the Wuhan Tovin Biotechnology Co., Ltd in China. ChIP-qPCR was used to confirm the direct binding of SB1 with its target genes and detect the acetylation level in sb1. The young panicles (< 3 cm) of SB1-GFP transgenic plants, sb1 mutant and wild type plants were collected for isolation of chromatin extracts. ChIP assays were performed using the EpiQuik™ Plant ChIP Kit (P-2014-48, Epigentek, USA), anti-GFP antibody (ab290, Abcam, USA) and anti-H3K9Ac antibody (ab10812, Abcam, USA). All qPCR experiments were conducted with the progress: 40 cycles of 95 ºC for 5 s, 60 ºC for 30 s and 72 ºC for 30 s in a reaction mixture containing 10 pmol of forward primer, 10 pmol of reverse primer and 1 μL of DNA from ChIP/control or 1 μL of input DNA diluted 20-fold (per biological replicate) as a template. More than three biological repeats with three technical repeats each were used for statistical analysis. The recovered DNA was used as the template for ChIP-qPCR with the method described previously (Zhuang et al, 2020). The primer sequences were listed in Table S2.
ACKNOWLEDGEMENTS
This study was supported by the National Natural Science Foundation of China (Grant No. 31971919), the National Key Program for Research and Development of China (Grant No. 2017YFD0100202), the Project Sponsored by Natural Science Foundation of Chongqing, China (Grant No. cstc2020jcyj- jqX0020) and Chongqing Graduate Research and Innovation Project funding in China (Grant No. CYS20123).
The transcription corepressor LEUNIG interacts with the histone deacetylase HDA19 and mediator components MED14 (SWP) and CDK8 (HEN3) to repress transcription
HuB, WangW, Ou SJ, Tang JY, LiH, Che RH, Zhang ZH, Chai XY, Wang HR, Wang YQ, Liang CZ, Liu LC, Piao ZZ, Deng QY, DengK, XuC, LiangY, Zhang LH, Li LG, Chu CC. 2015.
Variation in NRT1. 1B contributes to nitrate-use divergence between rice subspecies
Ikeda-KawakatsuK, MaekawaM, IzawaT, Itoh JI, NagatoY. 2012.
ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1
Overexpression of RCN1 and RCN2, rice TERMINAL FLOWER 1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice
WangW, HuB, Yuan DY, Liu YQ, Che RH, Hu YC, Ou SJ, Liu YX, Zhang ZH, Wang HR, LiH, Jiang ZM, Zhang ZL, Gao XK, Qiu YH, Meng XB, Liu YX, BaiY, LiangY, Wang YQ, Zhang LH, Li LG, SodmergenS, Jing HC, Li JY, Chu CC. 2018.
Expression of the nitrate transporter gene OsNRT1
.
1A/ OsNPF6.3 confers high yield and early maturation in rice
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
... To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
... In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010). ...
Inflorescence commitment and architecture in Arabidopsis
1
1997
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
LEUNIG, a putative transcriptional corepressor that regulates AGAMOUS expression during flower development
2
2000
... To illuminate the function of SB1, we carried out a yeast two- hybrid (Y2H) assay to find the interacting proteins of SB1. Firstly, the full-length SB1-BD plasmid was transformed into Y2HGold yeast cells. The yeast cells that harboured pGBKT7-FZP (positive control) can grow on both SD/-Trp and SD/-Trp-His-Ade media, whereas the yeast cells that harboured pGBKT7-SB1 and the empty pGBKT7 can only grow on the SD/-Trp medium but not on the SD/-Trp-His-Ade medium, indicating that SB1 protein showed no auto-activation ability (Fig. 5-A). Then, pGBKT7-SB1 was used as a bait to screen its partners in a cDNA library from rice young panicles. According to the screening results, we found two proteins: Os06g0126000 (OsSEUSS-like 1, OsSEU1) and Os04g0510200 (OsLEUNIG-like 1, OsLUG1) (Table S1), which are annotated as transcriptional corepressors and are homologous of SEU and LUG in Arabidopsis (http://rice.plantbiology. msu.edu/). Previous research had shown that the corepressor complex SEU-LUG plays an important role in plant development (Conner and Liu, 2000; Franks et al, 2002; Sridhar et al, 2004; Liu and Karmarkar, 2008; Sitaraman et al, 2008). Then, a Y2H assay was performed to test the interaction of SB1 with OsSEU1 and OsLUG1. As expected, after about 3 d, the yeast that co-expressed SB1 and OsLUG1 grew on the SD/-Trp-Leu-His-Ade selective medium, and the yeast that co-expressed SB1 and OsLUG1 grew after about 5 d. The results revealed that SB1 can interact with OsLUG1 and OsSEU1, and the interaction between SB1 and OsSEU1 was weaker than that of SB1 and OsLUG1 (Fig. 5-B). Next, a bimolecular fluorescent complimentary (BiFC) assay was conducted to further detect whether the SB1 can interact with SEU and LEU in leaf epidermal cells of Nicotiana benthamiana. Yellow fluorescent protein (YFP) signal was found in the nuclear of epidermal cells of tobacco leaves when co-expressing the SB1-YFPC fused protein with OsSEU1-YFPN and OsLUG1-YFPN fused proteins, respectively (Fig. 5-C). The results revealed that SB1 was capable of interaction with OsSEU1 and OsLUG1 in plant cells, and then acted as a transcriptional repressor in regulating the downstream genes. ...
... SB1 gene encodes a SHI/STY/SRS family protein and is allelic to OsSHI1. OsSHI1/SB1 gene regulates branch development in rice by acting as a repressive partner of IPA1 protein, a transcription activator of DEP1, to down-regulate the expression of DEP1 (Duan et al, 2019). In shi1, the mutant of complete- deletion of OsSHI1 gene, the numbers of secondary branches and spikelets increase sharply, and the DEP1 gene is also up-regulated significantly, similar with the sb1 mutants. In the present study, we showed a more precise mechanism of OsSHI1/SB1 regulating branch development. Firstly, the data of ChIP-seq and ChIP-qPCR showed SB1 can directly bind to the promoters of multiple panicle-related genes including TAW1, MOC1 and IPA1, besides DEP1. Secondly, we further revealed that SB1 can interact with two proteins of the SEU-LUG co-repressing complex, OsSEU1 and OsLUG1. SEU-LUG is known as a plant Gro/Tup1 co-repressor complex and responsible for transcriptional repressing in multiple development events. In Arabidopsis, the LUG-SEU complex with HDA19 can repress the AGMOUS (AG) expression in outer two whorls of floral organs via removing the acetylation mark from the AG locus (Conner and Liu, 2000; Liu and Karmarkar, 2008; Guo et al, 2015). Next, we also found the H3K9Ac level was increased in multiple sites of the ChIP-target genes. Therefore, our results together supported that SB1 can interact with the co-repressing complex OsSEU1-OsLUG1 and then directly repressed the expression of IPA1, DEP1, TAW1 and MOC1 through affecting the histone H3 acetylation level on target regions. In particular, the result supported that SB1 acted as a broad- spectrum transcriptional repressor on multiple targets in regulating panicle architecture in rice. Then, we speculated, it was the releasing of multiple branch development genes including IPA1, DEP1, TAW1 and MOC1 that eventually resulted in extremely prolonging BM fate and then dramatically increasing the numbers of spikelets in the sb1 and shi1 mutants (Fig. 7). ...
OsSHI1 regulates plant architecture through modulating the transcriptional activity of IPA1 in rice
1
2019
... SB1 gene encodes a SHI/STY/SRS family protein and is allelic to OsSHI1. OsSHI1/SB1 gene regulates branch development in rice by acting as a repressive partner of IPA1 protein, a transcription activator of DEP1, to down-regulate the expression of DEP1 (Duan et al, 2019). In shi1, the mutant of complete- deletion of OsSHI1 gene, the numbers of secondary branches and spikelets increase sharply, and the DEP1 gene is also up-regulated significantly, similar with the sb1 mutants. In the present study, we showed a more precise mechanism of OsSHI1/SB1 regulating branch development. Firstly, the data of ChIP-seq and ChIP-qPCR showed SB1 can directly bind to the promoters of multiple panicle-related genes including TAW1, MOC1 and IPA1, besides DEP1. Secondly, we further revealed that SB1 can interact with two proteins of the SEU-LUG co-repressing complex, OsSEU1 and OsLUG1. SEU-LUG is known as a plant Gro/Tup1 co-repressor complex and responsible for transcriptional repressing in multiple development events. In Arabidopsis, the LUG-SEU complex with HDA19 can repress the AGMOUS (AG) expression in outer two whorls of floral organs via removing the acetylation mark from the AG locus (Conner and Liu, 2000; Liu and Karmarkar, 2008; Guo et al, 2015). Next, we also found the H3K9Ac level was increased in multiple sites of the ChIP-target genes. Therefore, our results together supported that SB1 can interact with the co-repressing complex OsSEU1-OsLUG1 and then directly repressed the expression of IPA1, DEP1, TAW1 and MOC1 through affecting the histone H3 acetylation level on target regions. In particular, the result supported that SB1 acted as a broad- spectrum transcriptional repressor on multiple targets in regulating panicle architecture in rice. Then, we speculated, it was the releasing of multiple branch development genes including IPA1, DEP1, TAW1 and MOC1 that eventually resulted in extremely prolonging BM fate and then dramatically increasing the numbers of spikelets in the sb1 and shi1 mutants (Fig. 7). ...
SEUSS, a member of a novel family of plant regulatory proteins, represses floral homeotic gene expression with LEUNIG
1
2002
... To illuminate the function of SB1, we carried out a yeast two- hybrid (Y2H) assay to find the interacting proteins of SB1. Firstly, the full-length SB1-BD plasmid was transformed into Y2HGold yeast cells. The yeast cells that harboured pGBKT7-FZP (positive control) can grow on both SD/-Trp and SD/-Trp-His-Ade media, whereas the yeast cells that harboured pGBKT7-SB1 and the empty pGBKT7 can only grow on the SD/-Trp medium but not on the SD/-Trp-His-Ade medium, indicating that SB1 protein showed no auto-activation ability (Fig. 5-A). Then, pGBKT7-SB1 was used as a bait to screen its partners in a cDNA library from rice young panicles. According to the screening results, we found two proteins: Os06g0126000 (OsSEUSS-like 1, OsSEU1) and Os04g0510200 (OsLEUNIG-like 1, OsLUG1) (Table S1), which are annotated as transcriptional corepressors and are homologous of SEU and LUG in Arabidopsis (http://rice.plantbiology. msu.edu/). Previous research had shown that the corepressor complex SEU-LUG plays an important role in plant development (Conner and Liu, 2000; Franks et al, 2002; Sridhar et al, 2004; Liu and Karmarkar, 2008; Sitaraman et al, 2008). Then, a Y2H assay was performed to test the interaction of SB1 with OsSEU1 and OsLUG1. As expected, after about 3 d, the yeast that co-expressed SB1 and OsLUG1 grew on the SD/-Trp-Leu-His-Ade selective medium, and the yeast that co-expressed SB1 and OsLUG1 grew after about 5 d. The results revealed that SB1 can interact with OsLUG1 and OsSEU1, and the interaction between SB1 and OsSEU1 was weaker than that of SB1 and OsLUG1 (Fig. 5-B). Next, a bimolecular fluorescent complimentary (BiFC) assay was conducted to further detect whether the SB1 can interact with SEU and LEU in leaf epidermal cells of Nicotiana benthamiana. Yellow fluorescent protein (YFP) signal was found in the nuclear of epidermal cells of tobacco leaves when co-expressing the SB1-YFPC fused protein with OsSEU1-YFPN and OsLUG1-YFPN fused proteins, respectively (Fig. 5-C). The results revealed that SB1 was capable of interaction with OsSEU1 and OsLUG1 in plant cells, and then acted as a transcriptional repressor in regulating the downstream genes. ...
The Arabidopsis protein SHI represses gibberellin responses in Arabidopsis and barley
2
2001
... SB1 encodes a RING-like zinc finger transcription factor that belongs to SHI/LRP/SRS family. In Arabidopsis, the SHI family contains 10 members: SHORT INTERNODES (SHI), STYLISH1 and 2 (STY1 and STY2), SHI-RELATED SEQUENCE3 (SRS3) to SRS8, and LATERAL ROOT PRIMORDIUM (LRP) (Fridborg et al, 2001). Phylogenetic analysis revealed that there were five SHI/LRP/SRS protein family members in rice. SB1 had a poor homology with all SHI/LRP/SRS proteins in Arabidopsis and had a close evolutionary relationship with ZmLRL4 and ZmLRL5 in maize. ...
... The LOC_Os01g724901 and LOC_Os05g320701 had high homology and close branching relationship with LRP, which was widely studied in Arabidopsis and maize (Fig. 3-A). Previous studies have shown that the sequences of the SHI/LRP/SRS family proteins were highly diverse, with highly conserved regions: a C3HC3H RING-like domain and an IGGH domain (Fridborg et al, 2001; Kuusk et al, 2002, 2010). Then, the protein motif prediction program MEME and protein sequence alignment program DNAMAN were used to analyze the full-length amino acid sequences of SHI/LRP/SRS family proteins in rice, maize and Arabidopsis. The result showed that there were three highly conservative domain/motifs: a RING-like structure domain, an IGGH domain and an FRCVR domain in the SHI/LRP/SRS family proteins. In previous results, the FRCVR domain was not identified separately (Figs. 3-B, S2 and S3). ...
The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development
1
2010
... In this study, all the five mutants sb1-1/2/3/4/5 were manifested as extremely significant increase in the numbers of SBs and TBs (which barely appeared in the wild type plants), and finally showed an increase in the number of spikelets per panicle. Among them, the average ‘number of spikelets per panicle’ of sb1-1 reached nearly twice as much as the wild type. However, it was regretful that the number of grains per panicle in the sb1 mutant was obviously decreased because of the sharply decreased seed-setting rate. That meant the increasing numbers of spikelets in the sb1 mutant was meaningless unless handling the defect of seed-setting rate. In fact, it was reported that the advantageous traits (such as the increased number of branches) and disadvantageous factors (such as abnormal seed-setting rate and defect floral organs) for grain yield were concurrent in some mutants of panicle-related genes. For example, in the mutants of OsMADS34/PAP2, although the number of SBs increases sharply, the development of spikelet is affected and the seed-setting rate decreases (Gao et al, 2010; Kobayashi et al, 2010). Therefore, while the excellent natural variations in the genes such as Gn1a, TAW1, DEP1 and IPA1 had been used to largely benefit grain yield in rice, rebuilding and utilization of the genes pros and cons coexisting like SB1 and OsMADS34/PAP2 by the molecular design might be a future direction with quite development potentiality for the panicle architecture and grain yield in rice breeding. ...
The transcription corepressor LEUNIG interacts with the histone deacetylase HDA19 and mediator components MED14 (SWP) and CDK8 (HEN3) to repress transcription
1
2007
... Furthermore, it is reported that the LUG and SEU complex can interact with histone deacetylases to mediate the acetylation level of histone protein in the targeted region (Gonzalez et al, 2007; Liu and Karmarkar, 2008). We further detected the acetylation levels of histone protein H3 in the promoter and coding sequence region of DEP1, TAW1, MOC1 and IPA1 by ChIP-qPCR with an anti-H3K9Ac antibody. The result indicated that the acetylation levels of H3K9 in DEP1 (P1, P2 and P3 sites), TAW1 (P2 and P3 sites), MOC1 (P1, P4 and P5 sites) and IPA1 (P1 and P2 sites) were significantly increased in the sb1 mutant compared with those in the wild type (Fig. 6-D). These results suggested that SB1 regulated the transcription of DEP1, TAW1, MOC1 and IPA1 genes by the modification of histone acetylation. ...
Co-ordination of flower development through epigenetic regulation in two model species: Rice and Arabidopsis
1
2015
... SB1 gene encodes a SHI/STY/SRS family protein and is allelic to OsSHI1. OsSHI1/SB1 gene regulates branch development in rice by acting as a repressive partner of IPA1 protein, a transcription activator of DEP1, to down-regulate the expression of DEP1 (Duan et al, 2019). In shi1, the mutant of complete- deletion of OsSHI1 gene, the numbers of secondary branches and spikelets increase sharply, and the DEP1 gene is also up-regulated significantly, similar with the sb1 mutants. In the present study, we showed a more precise mechanism of OsSHI1/SB1 regulating branch development. Firstly, the data of ChIP-seq and ChIP-qPCR showed SB1 can directly bind to the promoters of multiple panicle-related genes including TAW1, MOC1 and IPA1, besides DEP1. Secondly, we further revealed that SB1 can interact with two proteins of the SEU-LUG co-repressing complex, OsSEU1 and OsLUG1. SEU-LUG is known as a plant Gro/Tup1 co-repressor complex and responsible for transcriptional repressing in multiple development events. In Arabidopsis, the LUG-SEU complex with HDA19 can repress the AGMOUS (AG) expression in outer two whorls of floral organs via removing the acetylation mark from the AG locus (Conner and Liu, 2000; Liu and Karmarkar, 2008; Guo et al, 2015). Next, we also found the H3K9Ac level was increased in multiple sites of the ChIP-target genes. Therefore, our results together supported that SB1 can interact with the co-repressing complex OsSEU1-OsLUG1 and then directly repressed the expression of IPA1, DEP1, TAW1 and MOC1 through affecting the histone H3 acetylation level on target regions. In particular, the result supported that SB1 acted as a broad- spectrum transcriptional repressor on multiple targets in regulating panicle architecture in rice. Then, we speculated, it was the releasing of multiple branch development genes including IPA1, DEP1, TAW1 and MOC1 that eventually resulted in extremely prolonging BM fate and then dramatically increasing the numbers of spikelets in the sb1 and shi1 mutants (Fig. 7). ...
Variation in NRT1. 1B contributes to nitrate-use divergence between rice subspecies
1
2015
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
Two splicing variants of OsNPF7.7 regulate shoot branching and nitrogen utilization efficiency in rice
1
2018
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
Natural variation at the DEP1 locus enhances grain yield in rice
3
2009
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
... In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010). ...
... In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets. ...
ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1
1
2012
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice
3
2010
... To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
... In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010). ...
... In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets. ...
PANICLE PHYTOMER2 (PAP2), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice
2
2010
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
... In this study, all the five mutants sb1-1/2/3/4/5 were manifested as extremely significant increase in the numbers of SBs and TBs (which barely appeared in the wild type plants), and finally showed an increase in the number of spikelets per panicle. Among them, the average ‘number of spikelets per panicle’ of sb1-1 reached nearly twice as much as the wild type. However, it was regretful that the number of grains per panicle in the sb1 mutant was obviously decreased because of the sharply decreased seed-setting rate. That meant the increasing numbers of spikelets in the sb1 mutant was meaningless unless handling the defect of seed-setting rate. In fact, it was reported that the advantageous traits (such as the increased number of branches) and disadvantageous factors (such as abnormal seed-setting rate and defect floral organs) for grain yield were concurrent in some mutants of panicle-related genes. For example, in the mutants of OsMADS34/PAP2, although the number of SBs increases sharply, the development of spikelet is affected and the seed-setting rate decreases (Gao et al, 2010; Kobayashi et al, 2010). Therefore, while the excellent natural variations in the genes such as Gn1a, TAW1, DEP1 and IPA1 had been used to largely benefit grain yield in rice, rebuilding and utilization of the genes pros and cons coexisting like SB1 and OsMADS34/PAP2 by the molecular design might be a future direction with quite development potentiality for the panicle architecture and grain yield in rice breeding. ...
Direct control of shoot meristem activity by a cytokinin-activating enzyme
1
2007
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
STY1 and STY2 promote the formation of apical tissues during Arabidopsis gynoecium development
1
2002
... The LOC_Os01g724901 and LOC_Os05g320701 had high homology and close branching relationship with LRP, which was widely studied in Arabidopsis and maize (Fig. 3-A). Previous studies have shown that the sequences of the SHI/LRP/SRS family proteins were highly diverse, with highly conserved regions: a C3HC3H RING-like domain and an IGGH domain (Fridborg et al, 2001; Kuusk et al, 2002, 2010). Then, the protein motif prediction program MEME and protein sequence alignment program DNAMAN were used to analyze the full-length amino acid sequences of SHI/LRP/SRS family proteins in rice, maize and Arabidopsis. The result showed that there were three highly conservative domain/motifs: a RING-like structure domain, an IGGH domain and an FRCVR domain in the SHI/LRP/SRS family proteins. In previous results, the FRCVR domain was not identified separately (Figs. 3-B, S2 and S3). ...
Functionally redundant SHI family genes regulate Arabidopsis gynoecium development in a dose-dependent manner
1
2010
... The LOC_Os01g724901 and LOC_Os05g320701 had high homology and close branching relationship with LRP, which was widely studied in Arabidopsis and maize (Fig. 3-A). Previous studies have shown that the sequences of the SHI/LRP/SRS family proteins were highly diverse, with highly conserved regions: a C3HC3H RING-like domain and an IGGH domain (Fridborg et al, 2001; Kuusk et al, 2002, 2010). Then, the protein motif prediction program MEME and protein sequence alignment program DNAMAN were used to analyze the full-length amino acid sequences of SHI/LRP/SRS family proteins in rice, maize and Arabidopsis. The result showed that there were three highly conservative domain/motifs: a RING-like structure domain, an IGGH domain and an FRCVR domain in the SHI/LRP/SRS family proteins. In previous results, the FRCVR domain was not identified separately (Figs. 3-B, S2 and S3). ...
Rice DENSE AND ERECT PANICLE 2 is essential for determining panicle outgrowth and elongation
2
2010
... To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
... ; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
Mutations in the F-box gene LARGER PANICLE improve the panicle architecture and enhance the grain yield in rice
2
2011
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
... In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010). ...
Short panicle1 encodes a putative PTR family transporter and determines rice panicle size
2
2009
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
... To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
Rice zinc finger protein DST enhances grain production through controlling Gn1a/OsCKX2 expression
1
2013
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
Identification and characterization of dwarf 62, a loss-of- function mutation in DLT/OsGRAS-32 affecting gibberellin metabolism in rice
2010
Control of tillering in rice
2
2003
... In rice, primary branches (PBs), secondary branches (SBs) and lateral spikelets (LSs) are all originated from axillary meristem, which compose the panicle with main axis and topic spikelet (TS) originating from apical meristem. Therefore, the genes participating in the initiation, formation and maintaining of axillary meristem in panicle are first regulators for panicle architecture and GNP, such as LAX PANICLE1 (LAX1), LAX2 and MONOCULM1 (MOC1) in rice. They all encode transcription factor, and the loss-of-function mutants of them all display sharp decreasing of branch and spikelet number (Li et al, 2003; Skirpan et al, 2008; Tabuchi et al, 2011). It is proved that LAX1 can interact with LAX2 and MOC1 respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development. ...
... In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets. ...
Genetic dissection of the source-sink relationship affecting fecundity and yield in rice (Oryza sativa L.)
1
1998
... In grass crops, grain number per panicle (GNP) is one of the most important yield components (Wang and Li, 2011). Panicle architecture consisting of the number of branches, the length of branches, and grain density on branches, determines the GNP (Li et al, 1998; Teng et al, 2002). In the past two decades, dozens of genes related to panicle architecture have been characterized in rice, some of which play crucial roles in determining GNP and then yield. ...
Groucho/Tup1 family co-repressors in plant development
3
2008
... To illuminate the function of SB1, we carried out a yeast two- hybrid (Y2H) assay to find the interacting proteins of SB1. Firstly, the full-length SB1-BD plasmid was transformed into Y2HGold yeast cells. The yeast cells that harboured pGBKT7-FZP (positive control) can grow on both SD/-Trp and SD/-Trp-His-Ade media, whereas the yeast cells that harboured pGBKT7-SB1 and the empty pGBKT7 can only grow on the SD/-Trp medium but not on the SD/-Trp-His-Ade medium, indicating that SB1 protein showed no auto-activation ability (Fig. 5-A). Then, pGBKT7-SB1 was used as a bait to screen its partners in a cDNA library from rice young panicles. According to the screening results, we found two proteins: Os06g0126000 (OsSEUSS-like 1, OsSEU1) and Os04g0510200 (OsLEUNIG-like 1, OsLUG1) (Table S1), which are annotated as transcriptional corepressors and are homologous of SEU and LUG in Arabidopsis (http://rice.plantbiology. msu.edu/). Previous research had shown that the corepressor complex SEU-LUG plays an important role in plant development (Conner and Liu, 2000; Franks et al, 2002; Sridhar et al, 2004; Liu and Karmarkar, 2008; Sitaraman et al, 2008). Then, a Y2H assay was performed to test the interaction of SB1 with OsSEU1 and OsLUG1. As expected, after about 3 d, the yeast that co-expressed SB1 and OsLUG1 grew on the SD/-Trp-Leu-His-Ade selective medium, and the yeast that co-expressed SB1 and OsLUG1 grew after about 5 d. The results revealed that SB1 can interact with OsLUG1 and OsSEU1, and the interaction between SB1 and OsSEU1 was weaker than that of SB1 and OsLUG1 (Fig. 5-B). Next, a bimolecular fluorescent complimentary (BiFC) assay was conducted to further detect whether the SB1 can interact with SEU and LEU in leaf epidermal cells of Nicotiana benthamiana. Yellow fluorescent protein (YFP) signal was found in the nuclear of epidermal cells of tobacco leaves when co-expressing the SB1-YFPC fused protein with OsSEU1-YFPN and OsLUG1-YFPN fused proteins, respectively (Fig. 5-C). The results revealed that SB1 was capable of interaction with OsSEU1 and OsLUG1 in plant cells, and then acted as a transcriptional repressor in regulating the downstream genes. ...
... Furthermore, it is reported that the LUG and SEU complex can interact with histone deacetylases to mediate the acetylation level of histone protein in the targeted region (Gonzalez et al, 2007; Liu and Karmarkar, 2008). We further detected the acetylation levels of histone protein H3 in the promoter and coding sequence region of DEP1, TAW1, MOC1 and IPA1 by ChIP-qPCR with an anti-H3K9Ac antibody. The result indicated that the acetylation levels of H3K9 in DEP1 (P1, P2 and P3 sites), TAW1 (P2 and P3 sites), MOC1 (P1, P4 and P5 sites) and IPA1 (P1 and P2 sites) were significantly increased in the sb1 mutant compared with those in the wild type (Fig. 6-D). These results suggested that SB1 regulated the transcription of DEP1, TAW1, MOC1 and IPA1 genes by the modification of histone acetylation. ...
... SB1 gene encodes a SHI/STY/SRS family protein and is allelic to OsSHI1. OsSHI1/SB1 gene regulates branch development in rice by acting as a repressive partner of IPA1 protein, a transcription activator of DEP1, to down-regulate the expression of DEP1 (Duan et al, 2019). In shi1, the mutant of complete- deletion of OsSHI1 gene, the numbers of secondary branches and spikelets increase sharply, and the DEP1 gene is also up-regulated significantly, similar with the sb1 mutants. In the present study, we showed a more precise mechanism of OsSHI1/SB1 regulating branch development. Firstly, the data of ChIP-seq and ChIP-qPCR showed SB1 can directly bind to the promoters of multiple panicle-related genes including TAW1, MOC1 and IPA1, besides DEP1. Secondly, we further revealed that SB1 can interact with two proteins of the SEU-LUG co-repressing complex, OsSEU1 and OsLUG1. SEU-LUG is known as a plant Gro/Tup1 co-repressor complex and responsible for transcriptional repressing in multiple development events. In Arabidopsis, the LUG-SEU complex with HDA19 can repress the AGMOUS (AG) expression in outer two whorls of floral organs via removing the acetylation mark from the AG locus (Conner and Liu, 2000; Liu and Karmarkar, 2008; Guo et al, 2015). Next, we also found the H3K9Ac level was increased in multiple sites of the ChIP-target genes. Therefore, our results together supported that SB1 can interact with the co-repressing complex OsSEU1-OsLUG1 and then directly repressed the expression of IPA1, DEP1, TAW1 and MOC1 through affecting the histone H3 acetylation level on target regions. In particular, the result supported that SB1 acted as a broad- spectrum transcriptional repressor on multiple targets in regulating panicle architecture in rice. Then, we speculated, it was the releasing of multiple branch development genes including IPA1, DEP1, TAW1 and MOC1 that eventually resulted in extremely prolonging BM fate and then dramatically increasing the numbers of spikelets in the sb1 and shi1 mutants (Fig. 7). ...
Genome-wide binding analysis of the transcription activator IDEAL PLANT ARCHITECTURE1 reveals a complex network regulating rice plant architecture
2
2013
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
... To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
Functional divergence of the TFL1-like gene family in Arabidopsis revealed by characterization of a novel homologue
1
2001
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
OsSPL14 promotes panicle branching and higher grain productivity in rice
2
2010
... In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010). ...
... In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets. ...
Overexpression of RCN1 and RCN2, rice TERMINAL FLOWER 1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice
2
2002
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
... In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets. ...
Distinct regulatory role for RFL, the rice LFY homolog, in determining flowering time and plant architecture
1
2008
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
Separation of shoot and floral identity in Arabidopsis
1
1999
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
A CONSTANS-like transcriptional activator, OsCOL13, functions as a negative regulator of flowering downstream of OsphyB and upstream of Ehd1 in rice
1
2016
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
Two-step regulation of a meristematic cell population acting in shoot branching in Arabidopsis
1
2016
... In rice, primary branches (PBs), secondary branches (SBs) and lateral spikelets (LSs) are all originated from axillary meristem, which compose the panicle with main axis and topic spikelet (TS) originating from apical meristem. Therefore, the genes participating in the initiation, formation and maintaining of axillary meristem in panicle are first regulators for panicle architecture and GNP, such as LAX PANICLE1 (LAX1), LAX2 and MONOCULM1 (MOC1) in rice. They all encode transcription factor, and the loss-of-function mutants of them all display sharp decreasing of branch and spikelet number (Li et al, 2003; Skirpan et al, 2008; Tabuchi et al, 2011). It is proved that LAX1 can interact with LAX2 and MOC1 respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development. ...
LEUNIG_HOMOLOG and LEUNIG perform partially redundant functions during Arabidopsis embryo and floral development
1
2008
... To illuminate the function of SB1, we carried out a yeast two- hybrid (Y2H) assay to find the interacting proteins of SB1. Firstly, the full-length SB1-BD plasmid was transformed into Y2HGold yeast cells. The yeast cells that harboured pGBKT7-FZP (positive control) can grow on both SD/-Trp and SD/-Trp-His-Ade media, whereas the yeast cells that harboured pGBKT7-SB1 and the empty pGBKT7 can only grow on the SD/-Trp medium but not on the SD/-Trp-His-Ade medium, indicating that SB1 protein showed no auto-activation ability (Fig. 5-A). Then, pGBKT7-SB1 was used as a bait to screen its partners in a cDNA library from rice young panicles. According to the screening results, we found two proteins: Os06g0126000 (OsSEUSS-like 1, OsSEU1) and Os04g0510200 (OsLEUNIG-like 1, OsLUG1) (Table S1), which are annotated as transcriptional corepressors and are homologous of SEU and LUG in Arabidopsis (http://rice.plantbiology. msu.edu/). Previous research had shown that the corepressor complex SEU-LUG plays an important role in plant development (Conner and Liu, 2000; Franks et al, 2002; Sridhar et al, 2004; Liu and Karmarkar, 2008; Sitaraman et al, 2008). Then, a Y2H assay was performed to test the interaction of SB1 with OsSEU1 and OsLUG1. As expected, after about 3 d, the yeast that co-expressed SB1 and OsLUG1 grew on the SD/-Trp-Leu-His-Ade selective medium, and the yeast that co-expressed SB1 and OsLUG1 grew after about 5 d. The results revealed that SB1 can interact with OsLUG1 and OsSEU1, and the interaction between SB1 and OsSEU1 was weaker than that of SB1 and OsLUG1 (Fig. 5-B). Next, a bimolecular fluorescent complimentary (BiFC) assay was conducted to further detect whether the SB1 can interact with SEU and LEU in leaf epidermal cells of Nicotiana benthamiana. Yellow fluorescent protein (YFP) signal was found in the nuclear of epidermal cells of tobacco leaves when co-expressing the SB1-YFPC fused protein with OsSEU1-YFPN and OsLUG1-YFPN fused proteins, respectively (Fig. 5-C). The results revealed that SB1 was capable of interaction with OsSEU1 and OsLUG1 in plant cells, and then acted as a transcriptional repressor in regulating the downstream genes. ...
Genetic and physical interaction suggest that BARREN STALK1 is a target of BARREN INFLORESCENCE2 in maize inflorescence development
1
2008
... In rice, primary branches (PBs), secondary branches (SBs) and lateral spikelets (LSs) are all originated from axillary meristem, which compose the panicle with main axis and topic spikelet (TS) originating from apical meristem. Therefore, the genes participating in the initiation, formation and maintaining of axillary meristem in panicle are first regulators for panicle architecture and GNP, such as LAX PANICLE1 (LAX1), LAX2 and MONOCULM1 (MOC1) in rice. They all encode transcription factor, and the loss-of-function mutants of them all display sharp decreasing of branch and spikelet number (Li et al, 2003; Skirpan et al, 2008; Tabuchi et al, 2011). It is proved that LAX1 can interact with LAX2 and MOC1 respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development. ...
Transcriptional repression of target genes by LEUNIG and SEUSS, two interacting regulatory proteins for Arabidopsis flower development
1
2004
... To illuminate the function of SB1, we carried out a yeast two- hybrid (Y2H) assay to find the interacting proteins of SB1. Firstly, the full-length SB1-BD plasmid was transformed into Y2HGold yeast cells. The yeast cells that harboured pGBKT7-FZP (positive control) can grow on both SD/-Trp and SD/-Trp-His-Ade media, whereas the yeast cells that harboured pGBKT7-SB1 and the empty pGBKT7 can only grow on the SD/-Trp medium but not on the SD/-Trp-His-Ade medium, indicating that SB1 protein showed no auto-activation ability (Fig. 5-A). Then, pGBKT7-SB1 was used as a bait to screen its partners in a cDNA library from rice young panicles. According to the screening results, we found two proteins: Os06g0126000 (OsSEUSS-like 1, OsSEU1) and Os04g0510200 (OsLEUNIG-like 1, OsLUG1) (Table S1), which are annotated as transcriptional corepressors and are homologous of SEU and LUG in Arabidopsis (http://rice.plantbiology. msu.edu/). Previous research had shown that the corepressor complex SEU-LUG plays an important role in plant development (Conner and Liu, 2000; Franks et al, 2002; Sridhar et al, 2004; Liu and Karmarkar, 2008; Sitaraman et al, 2008). Then, a Y2H assay was performed to test the interaction of SB1 with OsSEU1 and OsLUG1. As expected, after about 3 d, the yeast that co-expressed SB1 and OsLUG1 grew on the SD/-Trp-Leu-His-Ade selective medium, and the yeast that co-expressed SB1 and OsLUG1 grew after about 5 d. The results revealed that SB1 can interact with OsLUG1 and OsSEU1, and the interaction between SB1 and OsSEU1 was weaker than that of SB1 and OsLUG1 (Fig. 5-B). Next, a bimolecular fluorescent complimentary (BiFC) assay was conducted to further detect whether the SB1 can interact with SEU and LEU in leaf epidermal cells of Nicotiana benthamiana. Yellow fluorescent protein (YFP) signal was found in the nuclear of epidermal cells of tobacco leaves when co-expressing the SB1-YFPC fused protein with OsSEU1-YFPN and OsLUG1-YFPN fused proteins, respectively (Fig. 5-C). The results revealed that SB1 was capable of interaction with OsSEU1 and OsLUG1 in plant cells, and then acted as a transcriptional repressor in regulating the downstream genes. ...
LAX PANICLE2 of rice encodes a novel nuclear protein and regulates the formation of axillary meristems
2
2011
... In rice, primary branches (PBs), secondary branches (SBs) and lateral spikelets (LSs) are all originated from axillary meristem, which compose the panicle with main axis and topic spikelet (TS) originating from apical meristem. Therefore, the genes participating in the initiation, formation and maintaining of axillary meristem in panicle are first regulators for panicle architecture and GNP, such as LAX PANICLE1 (LAX1), LAX2 and MONOCULM1 (MOC1) in rice. They all encode transcription factor, and the loss-of-function mutants of them all display sharp decreasing of branch and spikelet number (Li et al, 2003; Skirpan et al, 2008; Tabuchi et al, 2011). It is proved that LAX1 can interact with LAX2 and MOC1 respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development. ...
... respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development. ...
MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods
1
2011
... The SB1-related protein sequences were downloaded from GenBank (http://www.ncbi.nlm.nih.gov/genbank/) using the full-length amino acid sequence of SB1 as a query. A phylogenetic tree was constructed using the maximum likelihood method based on the Jones-Taylor-Thornton matrix-based model with the lowest Bayesian information criterion scores by MEGA 5.0 (Tamura et al, 2011). Statistical support for the tree topology was assessed by means of a bootstrap analysis with 500 replicates. Protein motifs were predicted using the motif-based sequence analysis tool MEME (http://alternate.meme-suite.org/). ...
QTL analysis of rice peduncle vascular bundle system and panicle traits
1
2002
... In grass crops, grain number per panicle (GNP) is one of the most important yield components (Wang and Li, 2011). Panicle architecture consisting of the number of branches, the length of branches, and grain density on branches, determines the GNP (Li et al, 1998; Teng et al, 2002). In the past two decades, dozens of genes related to panicle architecture have been characterized in rice, some of which play crucial roles in determining GNP and then yield. ...
Ectopic expression of OsMADS1 caused dwarfism and spikelet alteration in rice
1
2017
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
1A/ OsNPF6.3 confers high yield and early maturation in rice
1
2018
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
Branching in rice
1
2011
... In grass crops, grain number per panicle (GNP) is one of the most important yield components (Wang and Li, 2011). Panicle architecture consisting of the number of branches, the length of branches, and grain density on branches, determines the GNP (Li et al, 1998; Teng et al, 2002). In the past two decades, dozens of genes related to panicle architecture have been characterized in rice, some of which play crucial roles in determining GNP and then yield. ...
DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously
1
2010
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
The QTL GNP1 encodes GA20ox1, which increases grain number and yield by increasing cytokinin activity in rice panicle meristems
1
2016
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
STAMENLESS 1, encoding a single C2H2 zinc finger protein, regulates floral organ identity in rice
1
2009
... To construct a complementary plasmid, a 4091-bp genomic fragment that contained the 3070-bp upstream sequence and 1021-bp SB1 coding sequence coupled with GFP reporter gene coding sequence was amplified using the primers SB1-comF and SB1-comR (Table S2). The fragment was inserted into a binary vector pCAMBIA1300 using the pEASY®-Uni Seamless Cloning and Assembly Kit (TRANSGENE, Beijing, China) to construct the pCAMBIA1300-SB1-GFP vector. The recombinant plasmids were transformed into the sb1 mutant using the Agrobacterium tumefaciens-mediated transformation method as described previously (Xiao et al, 2009). ...
Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice
2
2008
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
... To sum up, the genes related to GNP are involved in multiple networks and they have many members, some of which have been used to improve GNP and thus yield in the long period of productive practice, such as SP1, DEP1, IPA1, Gn1a and TWA1 (Ashikari et al, 2005; Xue et al, 2008; Li et al, 2009; Jiao et al, 2010; Li F et al, 2010; Li W Q et al, 2010; Lu et al, 2013). However, as the exhausted potential of old genes, there is an urgent need to characterize new genes related to panicle architecture and GNP in rice. In the present study, we identified five allelic mutants, which were characterized by significant increase of the numbers of branches and spikelets, and named super branch 1 (sb1). SB1 gene encodes a new RING- like zinc finger protein, locates in nuclear and is expressed highly in BM. By analysis of qPCR and chromatin immunoprecipitation (ChIP) assays, we found SB1 participates in direct repressing multiple known genes that related to panicle development, including IPA1, DEP1, TAW1 and MOC1. Therefore, our data implied that SB1 acts as a transcription repressor and functions on accelerating the transition of BM to SM in panicle architecture of rice. ...
A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice
1
2011
... Some genes which are involved in heading date and transporting nutrients can also control panicle architecture, maybe because of the delayed heading date or increased nitrate in panicles prolonging the BM activity. Grains.Height.Date-7 (Ghd7) encodes a CCT-domain protein, high expression of which greatly increases GNP and the overall yield per plant under long-day conditions, while Ghd8 and OsCOL13 have similar effects (Xue et al, 2008; Wei et al, 2010; Yan et al, 2011; Sheng et al, 2016). Nitrate transporter 1/peptide transporter (NPF) family members (NPF2.2, NPF4.1/ sp1, NPF6.3, NPF7.3 and NPF7.7) have been proved to regulate panicle branching by transporting nitrate or other unknown nutrients. Overexpression of most of them can increase panicle branch number by enhancing nitrogen utilization (Li et al, 2009; Hu et al, 2015; Huang et al, 2018; Wang et al, 2018). ...
Chromatin interacting factor OsVIL2 increases biomass and rice grain yield
1
2019
... Extensive research has demonstrated that plant hormones play crucial roles in panicle architecture, such as cytokinin. Grain number 1a (Gn1a), a major QTL of GNP, encodes cytokinin oxidase 2 (OsCKX2) which is responsible for cytokinin degradation. In the natural allelic variant with low-level expression of Gn1a, the accumulation of cytokinin leads to largely increased branch and spikelet numbers and thus grain yield (Ashikari et al, 2005). It has been confirmed that multiple genes participate in panicle development by regulating the expression of Gn1a. The zinc finger transcription factor DROUGHT AND SALT TOLERANCE (DST) activates Gn1a expression by directly binding to its promoter (Li et al, 2013), while the chromatin interacting factor VIN3-LIKE 2 (VIL2) directly restricts Gn1a expression by binding with its promoter region (Yang et al, 2019). When the loss-of-function of DST results in low-level Gn1a expression and then increases branches, overexpression of VIL2 leads to increased branches as low expression of Gn1a. The microRNA156-IPA1-DEP1 pathway regulates panicle development through cytokinin pathway, in which IDEAL PLANT ARCHITECTUTRE1 (IPA1) (OsSPL14/ WEP) can directly activate DENSE AND ERECT PANICLE1 (DEP1) expression by binding with its promoter, and then high expression of DEP1 represses Gn1a (Huang et al, 2009; Lu et al, 2013). The mutation of LARGER PANICLE (LP) increased panicle branching by down-regulating the Gn1a transcript (Li et al, 2011). In addition, LONELY GUY (LOG) (a cytokinin- activating enzyme), Grain Number per Panicle1 (GNP1, the gibberellin oxidase GA20ox1) and transcript activator SP3 regulate branch development by affecting cytokinin synthesis (Kurakawa et al, 2007; Wu et al, 2016). ...
TAWAWA1, a regulator of rice inflorescence architecture, functions through the suppression of meristem phase transition
3
2013
... The transition from branch meristem (BM) to spikelet meristem (SM) is also considered as a key process for panicle architecture. The delay of the transition can elongate the BM fate and then results in more and longer branches, and then more spikelets, whereas the earlier transition results in less spikelets. In Arabidopsis, the mutual restriction of TERMINAL FLOWER 1 (TFL1, an inflorescence meristem identity gene) and LEAFY (LFY, a floral organ identity gene) determines the inflorescence architecture (Bradley et al, 1997; Ratcliffe et al, 1999; Mimida et al, 2001). RICE CENTRORADIALIS1 (RCN1) and RCN2, two orthologs of TFL1, maintain the BM fate in rice, overexpression of which leads to the delayed transition from BM to SM, and then more branches and spikelets (Nakagawa et al, 2002). However, ABERRANT PANICLE ORGANIZATION 2 (APO2)/RFL, functions on BM but not SM in rice, an opposite effect with its ortholog LFY. It is also proved that APO2/RFL can be activated by a DOF-domain transcription activator SHORT PANICLE 3 (SP3), and interact with APO1, to delay the transition from BM to SM. In mutants of apo1, apo2 and sp3, the numbers of branches and spikelets are all decreased significantly (Rao et al, 2008; Ikeda- Kawakatsu et al, 2012). In addition, TAWAWA1 (TAW1) encodes a nuclear protein and also participates in determining the fate of BM. In gain-of-function mutant taw1-D, the BM activity is increased and the differentiation of SM is delayed, which improves the formation of more SBs and spikelets (Yoshida et al, 2013). Two E-class MADS-box factors, OsMADS1 and OsMADS34/PAP2, might accelerate the differentiation of SM. Overexpression of OsMADS1 leads to the less numbers of branches and spikelets, whereas the pap2 mutant (one of the loss-of-function of OsMADS34) displays the more numbers of branches and spikelets (Kobayashi et al, 2010; Wang et al, 2017). ...
... In rice, GNP as one of three key factors of grain yield, is heavily influenced by the inflorescence architecture, especially the number of branches. The previous research showed that a large number of genes can affect GNP to increase/decrease the yield by regulating the number of branches. Mutations of Gn1a, TAW1, DEP1, LP and IPA1 all lead to increased branches and GNP to finally increase the yield. Gn1a gene acts as a negative regulator of the yield by degrading the cytokinin. Mutation of Gn1a increases the number of spikelets by 20% and significantly increases the yield of rice (Ashikari et al, 2005). In the gain-functional mutant tawawa1-D, the spikelet specialization is delayed, resulting in prolonged branch formation time, increased the numbers of SBs and spikelets per panicle and finally the yield (Yoshida et al, 2013). The functional acquired mutation of DEP1 makes rice panicle dense and increases the numbers of branches and grains per panicle, thus promoting rice yield of 15%‒20% (Huang et al, 2009). LP gene encodes an F-box protein, and its mutation leads to a significant increase in the numbers of branches and spikelets, with a yield increase of about 11% per plant (Li et al, 2011). QTL IPA1 is an allele of OsSPL14. Mutation of OsSPL14 disturbs OsmiR156’s regulation of OsSPL14, resulting in reduced tillering and significantly increasing branch number, and then panicle grain number and the yield by 10% (Jiao et al, 2010; Miura et al, 2010). ...
... In the sb1 panicles, as more SBs and TBs were produced, the expression of some genes related to initiate and maintain BM fate was up-regulated, such as RCN1, MOC1, MOC3, TAW1, IPA1 and DEP1. It was reported that enhancing the function of most of these genes can promote branch development in rice (Nakagawa et al, 2002; Li et al, 2003; Huang et al, 2009; Jiao et al, 2010; Miura et al, 2010; Yoshida et al, 2013). These results together suggested that SB1 might be responsible for accelerating the transition from BM to SM in panicles, and the mutations of SB1 prolonged the BM fate and then resulted in more secondary and tertiary branches instead of producing spikelets. ...
LATERAL FLORET 1 induced the three-florets spikelet in rice
1
2017
... In rice, primary branches (PBs), secondary branches (SBs) and lateral spikelets (LSs) are all originated from axillary meristem, which compose the panicle with main axis and topic spikelet (TS) originating from apical meristem. Therefore, the genes participating in the initiation, formation and maintaining of axillary meristem in panicle are first regulators for panicle architecture and GNP, such as LAX PANICLE1 (LAX1), LAX2 and MONOCULM1 (MOC1) in rice. They all encode transcription factor, and the loss-of-function mutants of them all display sharp decreasing of branch and spikelet number (Li et al, 2003; Skirpan et al, 2008; Tabuchi et al, 2011). It is proved that LAX1 can interact with LAX2 and MOC1 respectively to regulate the development of axillary meristem (Tabuchi et al, 2011), but the targets of them are still unclear up to date. In Arabidopsis, LATERAL SUPPRESSOR (LAS) (an ortholog of MOC1) can directly regulate REVOLUTA (REV, a HD-ZIPIII protein), which can activate meristem maintaining gene SHOOT MERISTEMLESS (STM), to promote the initiation and formation of axillary meristem (Shi et al, 2016). LATERAL FLORET 1 (LF1, an ortholog of REV in rice) regulates lateral floret formation in rice spikelets by activating OSH1 (an ortholog of STM in rice) (Zhang et al, 2017). However, a similar pathway of MOC1-LF1-OSH1 has not been proved in branch formation in rice panicle development. ...
NONSTOP GLUMES1 encodes a C2H2 zinc finger protein that regulates spikelet development in rice
1
2020
... The young panicles (< 3 cm) from the SB1P:SB1:GFP complementary transgenic plants were collected to do the ChIP-seq using a GFP-antibody. The sequencing and data analysis were carried out by the Wuhan Tovin Biotechnology Co., Ltd in China. ChIP-qPCR was used to confirm the direct binding of SB1 with its target genes and detect the acetylation level in sb1. The young panicles (< 3 cm) of SB1-GFP transgenic plants, sb1 mutant and wild type plants were collected for isolation of chromatin extracts. ChIP assays were performed using the EpiQuik™ Plant ChIP Kit (P-2014-48, Epigentek, USA), anti-GFP antibody (ab290, Abcam, USA) and anti-H3K9Ac antibody (ab10812, Abcam, USA). All qPCR experiments were conducted with the progress: 40 cycles of 95 ºC for 5 s, 60 ºC for 30 s and 72 ºC for 30 s in a reaction mixture containing 10 pmol of forward primer, 10 pmol of reverse primer and 1 μL of DNA from ChIP/control or 1 μL of input DNA diluted 20-fold (per biological replicate) as a template. More than three biological repeats with three technical repeats each were used for statistical analysis. The recovered DNA was used as the template for ChIP-qPCR with the method described previously (Zhuang et al, 2020). The primer sequences were listed in Table S2. ...
 , Yunfeng Li
, Yunfeng Li
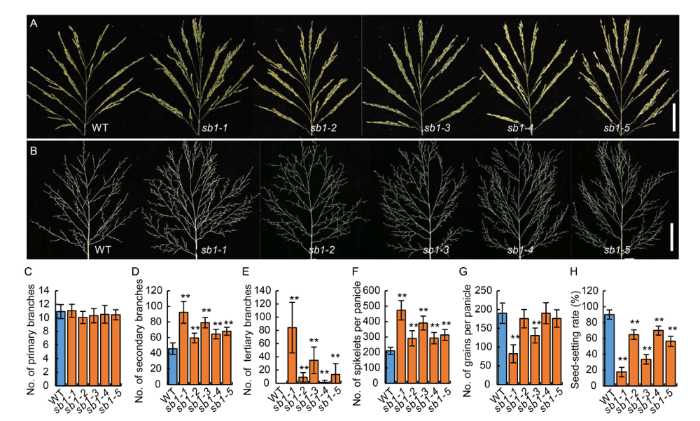
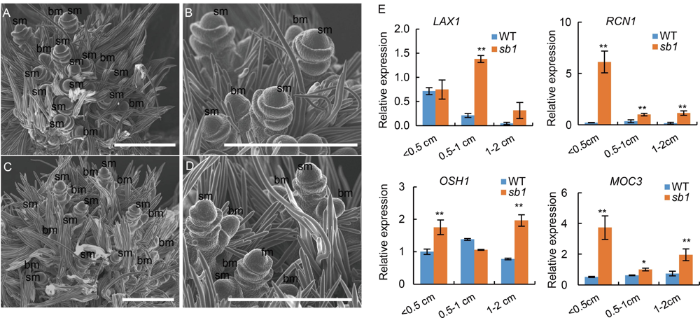
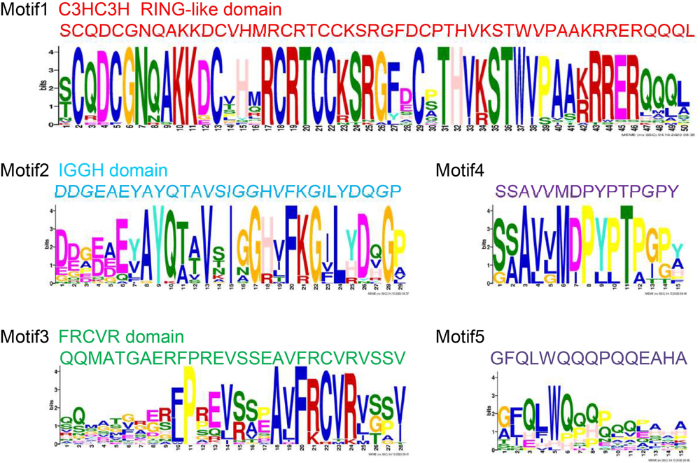

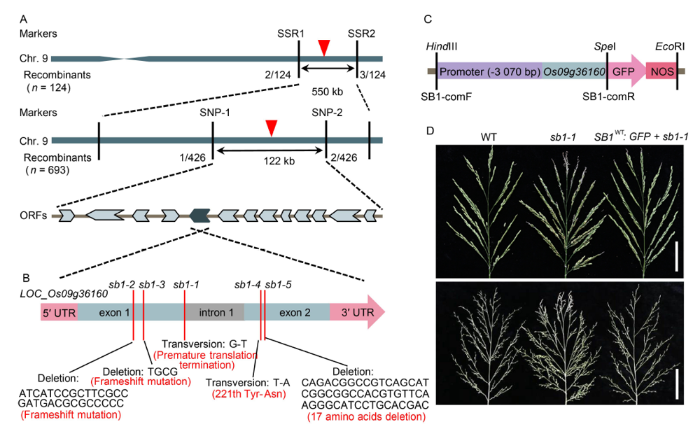
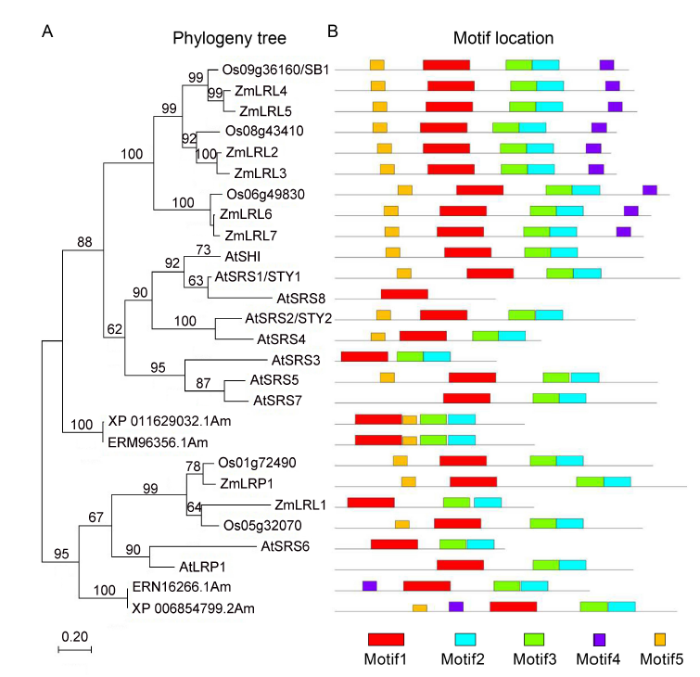
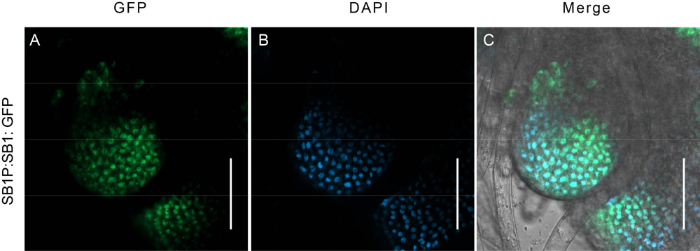
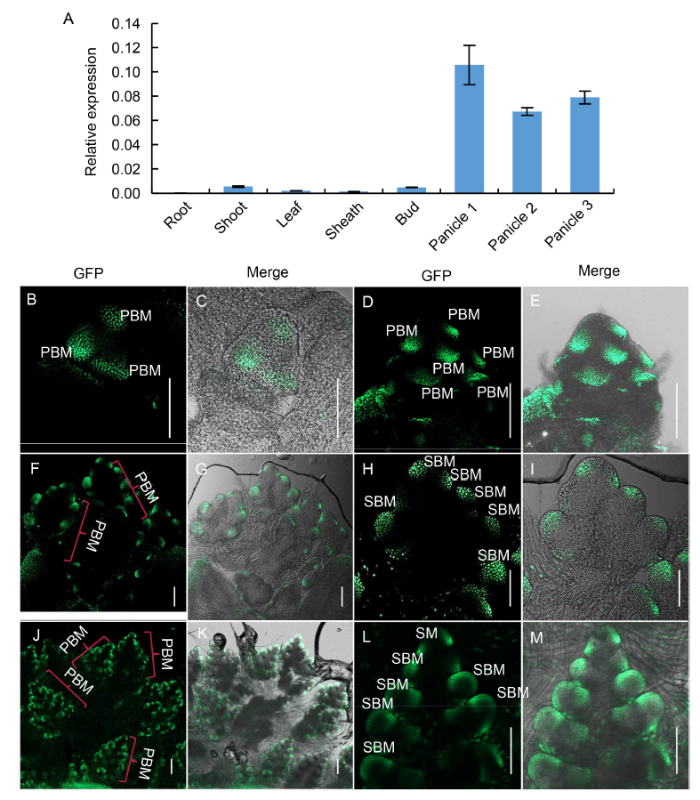
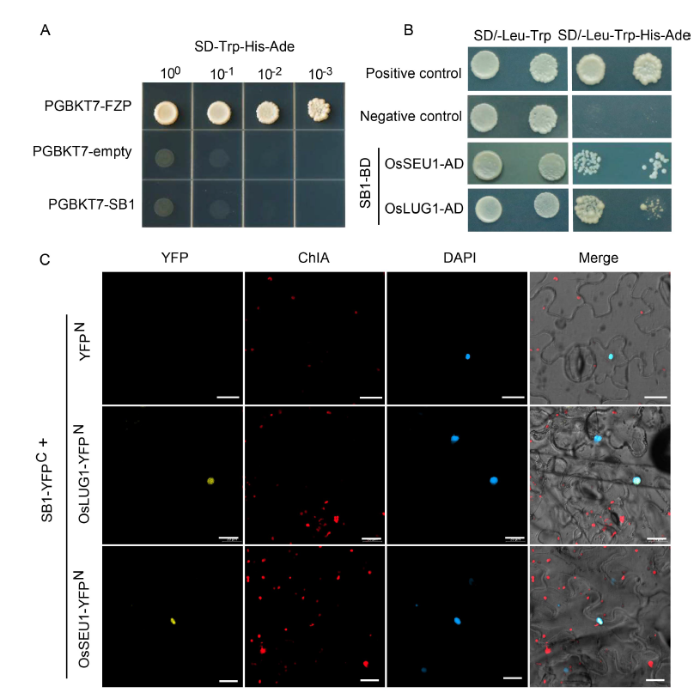
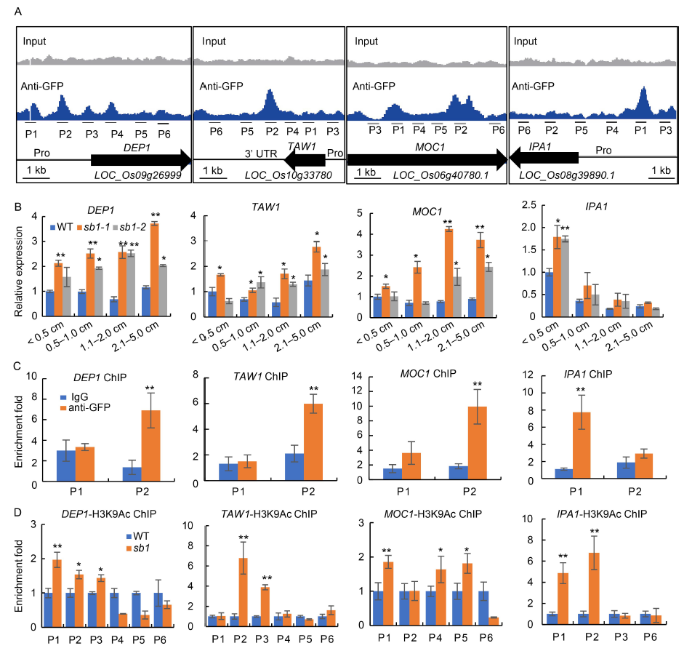
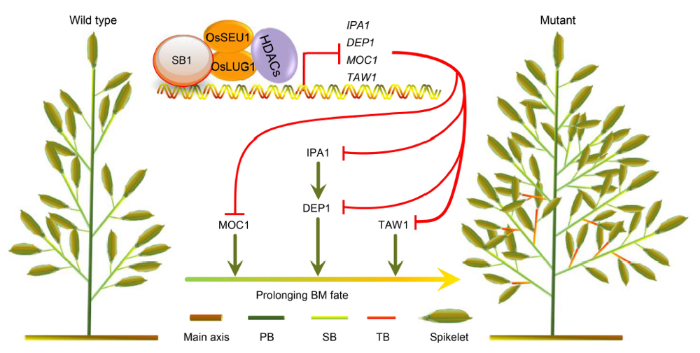
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}